草原旅人的自然誌:從西伯利亞到台灣田野的長途遷徙者
第一章 物種概述:基本資訊與分類
在台灣冬季的田野邊緣,偶爾可以看見一隻身形修長的褐色小鳥,從乾燥的草叢中拔腿直立,以一種幾乎令人訝異的筆直姿態站在泥地上,兩腿細長,身體比例較其他常見麻雀、鶺鴒更為高挑。牠在原地停頓片刻,以圓大的眼睛掃視四周,然後緩步向前,每一步之間都有一個細微的尾部上下抽動動作。這種鳥不如白鶺鴒那樣顯眼,也不如燕雀那樣常見,甚至在許多賞鳥者拿起望遠鏡仔細核對辨識特徵之前,往往會被粗略歸入「又一隻鷚」的模糊印象中,一眼帶過。然而,這個沉默的草地旅人擁有一段跨越六千公里的生命旅程,以及一個在分類學史上橫跨近兩百年、涉及超過十個物種名稱的複雜命名故事——牠,就是大花鷚(Anthus richardi)。
1.1 中文名、俗名與多語言名號的命名脈絡
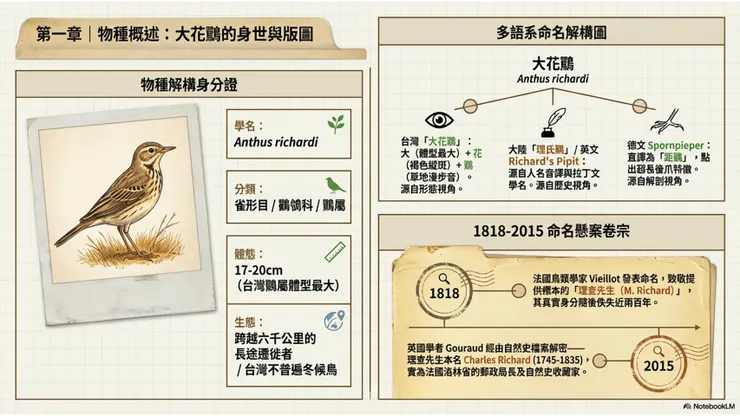
大花鷚的中文正式名稱「大花鷚」,由兩個辨識層次的詞根組成:「大」字標示其在鷚ㄐㄩㄣˋ(jùn)屬(Anthus)已知台灣記錄物種中體型最大的相對位置;「花」字描述其褐色底羽上遍布深褐色縱斑的斑紋羽衣,如同印在布料上的花紋;「鷚」則是中文鳥類命名系統中對鶺鴒科鷚屬物種的通用字,音「溜(liù)」,象聲兼會意,既摹擬鷚類在草地上行走時細小輕快的步音,也暗示其棲息於開闊草地、低矮植被環境的生態習性。三字合而為「花紋繁複的大型鷚鳥」,是一個準確而簡練的描述性命名。
在中國大陸的鳥類文獻系統中,本種通用名為「理氏鷚(Lǐ shì liù)」,是英文學名 richardi 的音譯,取「理查(Richard)」的中文音近譯字「理氏」,加上「鷚」字組合而成;這一命名直接對應英文名「Richard's Pipit」的命名邏輯,清楚地記錄了此種以人名命名的歷史。台灣中文名「大花鷚」與大陸「理氏鷚」的並行,反映了兩岸在鳥類中文命名系統上長期平行發展所形成的差異——台灣傾向以形態描述命名,大陸則更常採用音譯人名命名,兩種命名邏輯各有其文化背景與使用慣性,在查閱資料時必須注意這一差異以避免混淆。此外,本種在台灣早期文獻中偶見「田鷚」的舊稱,強調其田野棲息習性,但此名已逐漸少用,現行台灣鳥類名錄以「大花鷚」為正式通用名。
多語言命名方面,英文「Richard's Pipit」是全球最通行的稱呼,另有「Oriental Pipit」(東方鷚)的舊稱,後者是因其分布於東方古北界而得名,但「Oriental Pipit」這一名稱因曾被用於多個不同的鷚屬物種,造成混淆,在現代文獻中已逐漸廢棄不用。德文「Spornpieper」(距鷚)直接點出本種最重要的形態特徵——後趾的超長距爪(hindclaw,即「距」),以此作為命名核心,與英文的人名命名策略形成有趣對比。法文「Pipit de Richard」、西班牙文「Bisbita de Richard」均採用人名加鷚類通稱的命名模式,與英文保持一致。日文「ムネアカタヒバリ(Mune-aka-tahibari)」中,「ムネアカ」意為「胸紅」,然而此日文名其實對應的是赤喉鷚(Anthus cervinus)而非大花鷚,大花鷚在日文中的正確學名對應名稱應以學名核查為準,這一點在使用日文資料時需特別留意。泰文「นกเด้าดินทุ่งใหญ่」(nok-dao-din-thung-yai)意為「大草地地鳥」,以體型(大)、棲地(草地)和地面活動習性(地鳥)三個特徵組合命名,是一個頗為實用的描述性命名。
1.2 學名 Anthus richardi 的語源、命名者維埃約與「理查先生」的身份謎案
大花鷚的學名 Anthus richardi 包含兩個部分:屬名 Anthus 源自拉丁文,本是古羅馬博物學家用於指稱一種棲息於草地、習慣在牛蹄旁覓食小蟲的小型鳥類的詞彙,其詞源可能來自希臘文,意指「草地上的鳥」;這個屬名由法國植物學家安托萬·洛朗·德·裘西厄(Antoine Laurent de Jussieu)確立為鷚屬的學名,精準地概括了整個屬的棲地特徵。種小名 richardi 則是一個拉丁文屬格(genitive case),意為「屬於理查(Richard)的」,以紀念為這種鳥的學術命名提供標本的重要人物。
命名者路易·讓—皮耶·維埃約(Louis Jean Pierre Vieillot,1748—1831)是法國著名鳥類學家,也是19世紀初歐洲最多產的鳥類分類學者之一,其一生描述命名的鳥類物種及亞種數量超過數百個,對近代鳥類系統分類學的建立貢獻卓著。維埃約於1818年在其編撰的《應用於藝術農業及醫學的新自然史詞典》(Nouveau Dictionnaire d'Histoire Naturelle Appliquée aux Arts)第二十六卷第491頁,以「Le Pipi Richard,Anthus Richardi」的名稱正式發表了大花鷚,命名所依據的是三件採自法國東北部洛林(Lorraine)地區的同模標本(syntypes)。在命名說明中,維埃約明確指出,這個物種是以「理查先生(M. Richard de Lunéville)」命名的,「他使我知悉此鳥,並使我得以在新版詞典中加以描述」。
然而,在其後近兩百年的鳥類學文獻史中,「理查先生」的真實身份幾乎完全消失在歷史的記載之中,成為鳥類命名史上的一個懸案。直至2015年,英國鳥類學家克里斯托弗·古拉德(Christophe Gouraud)在《自然史檔案》(Archives of Natural History)期刊發表的研究論文中,終於通過翻查洛林省的歷史檔案和自然史收藏記錄,確認了「理查先生」的真實身份:此人為夏爾·理查(Charles Richard,1745—1835),時任法國洛林省呂內維爾(Lunéville)的郵政局長(director of the postal service),同時是著名南非博物學家弗朗索瓦·勒瓦揚(François Levaillant)的私人好友,以及當時頗負盛名的自然史標本收藏者。理查收藏的部分標本現仍存放於法國拉沙特爾(La Châtre)的貝永(Baillon)收藏館中,但包括大花鷚三件同模標本在內的多件標本已下落不明,無從追溯。
這段命名歷史提供了一個關於科學命名實踐的微觀案例:物種的學名常常是對提供標本、採集信息或提供資助的當地收藏家的紀念,但這些被紀念的人物在其後的歷史中往往因缺乏獨立的科學著述而逐漸被遺忘,最終只留下一個拉丁文屬格後綴,懸掛在物種名稱之後,等待好奇的研究者去重新發掘他們的真實故事。
1.3 分類地位:鶺鴒科鷚屬的系統位置
大花鷚在生物系統分類學中的位置為:脊索動物門(Chordata)、脊椎動物亞門(Vertebrata)、鳥綱(Aves)、雀形目(Passeriformes)、鶺鴒科(Motacillidae)、鷚屬(Anthus)。鶺鴒科(Motacillidae)是雀形目中一個主要分布於舊世界的科,全科包含鷚屬(Anthus)、鶺鴒屬(Motacilla)、長爪鷚屬(Macronyx)及山鶺鴒屬(Dendronanthus)等數個屬,約65至68個物種(依不同分類系統而略有差異),廣泛分布於全球各洲(南極洲除外),以非洲和亞洲的多樣性最高。
鷚屬(Anthus)是鶺鴒科中物種數量最多的屬,現行分類系統(IOC世界鳥類名錄)認定約有40至43個有效種,分布範圍涵蓋全球各大洲,從北極凍原到熱帶草原,從高山草甸到海濱礫灘,各類開闊的非林地環境中均有鷚屬物種的蹤跡。鷚屬物種的共同特徵包括:體型中小型(體長14至22公分)、體色以褐色縱斑為主(開闊地保護色)、地棲性強(主要在地面行走覓食)、尾部抽動習慣(tail-wagging),以及後趾具有較長後爪(hindclaw)等。在鶺鴒科的系統位置方面,分子系統學研究顯示鶺鴒科在雀形目中屬於「鴉雀總科(Passeroidea)」大支系,與鶲科(Muscicapidae)、文鳥科(Estrildidae)等科存在不同程度的親緣關係。
在台灣的記錄中,鷚屬共有六個物種具有確認的野外記錄,依常見程度依次為:樹鷚(Anthus hodgsoni,普遍冬候鳥)、赤喉鷚(Anthus cervinus,普遍冬候鳥)、大花鷚(Anthus richardi,不普遍冬候鳥)、白背鷚(Anthus gustavi,不普遍過境鳥)、布萊氏鷚(Anthus godlewskii,罕見迷鳥)及稻田鷚(Anthus rufulus,罕見迷鳥)。大花鷚在這六種台灣鷚屬記錄中,以體型最大、後爪最長,以及叫聲最具爆發力而最具辨識度,但其「不普遍冬候鳥」的定性,意味著在台灣各地的年度出現頻率和族群密度,均不及樹鷚和赤喉鷚那般穩定常見,是需要特意尋訪才有機會見到的物種。
1.4 歷史分類沿革:從 Anthus novaeseelandiae 超種複合體到獨立物種的分裂之路
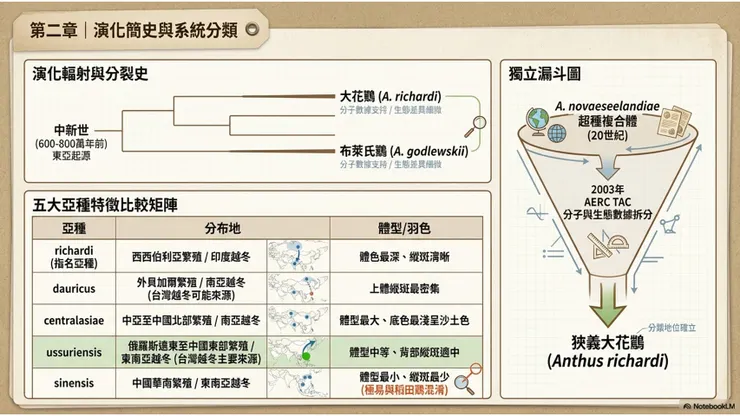
大花鷚的分類學歷史是一段漫長而複雜的「合合分分」過程,至今仍是鳥類系統學文獻中爭議持續的議題之一。在20世紀的大部分時間裡,大花鷚、澳大拉西亞鷚(Anthus novaeseelandiae)、非洲鷚(African Pipit)、山鷚(Mountain Pipit)和稻田鷚(Anthus rufulus)等形態高度相似的鷚屬物種,被多數分類系統合併處理為同一個廣義物種「大花鷚(Anthus novaeseelandiae)」的不同亞種群,形成一個所謂的「超種複合體(superspecies complex)」。在這一分類框架下,「大花鷚」這個名稱本身指涉的實際上是一個涵蓋從新西蘭到非洲草原、再到西伯利亞草地的廣大地理範圍內多個形態相似族群的集合概念,而非一個嚴格定義的物種個體。
這一寬鬆的廣義物種概念在20世紀末開始受到挑戰。隨著分子系統學技術的應用以及對各族群繁殖生物學、叫聲特徵和羽色形態的深入比較,各獨立研究相繼指出,這些被合併處理的族群在遺傳、叫聲和形態上均存在足以支持物種獨立地位的差異。歐洲鳥類記錄委員會分類顧問委員會(AERC TAC)在2003年正式建議,將原「A. novaeseelandiae 超種複合體」拆分為五個獨立物種,其中以維埃約1818年命名的指名亞種 richardi 為代表的廣義大花鷚(包含 richardi、dauricus、centralasiae、ussuriensis、sinensis 等亞種)被確立為獨立物種,恢復使用 Anthus richardi 的學名。這一建議在其後逐漸被IOC世界鳥類名錄、HBW鳥類手冊及多數現行分類系統所接受,大花鷚作為獨立物種的分類地位逐漸確立。
然而,分裂的爭議並未就此終止。稻田鷚(Anthus rufulus)與大花鷚的種界在部分東南亞地區仍存在重疊的中間型個體,使得在越冬地區(包括台灣偶有的稻田鷚記錄)進行精確的物種識別時需要特別謹慎;非洲鷚的分類地位在部分系統中仍有爭議;而與大花鷚親緣最近的布萊氏鷚(Anthus godlewskii)在台灣目前被列為迷鳥,但其與大花鷚在形態上的高度相似性,使得每逢冬季在台灣農田出現體型較大的鷚類時,精確辨識的挑戰始終存在。這一系列仍在進行中的分類學討論,使大花鷚成為理解「物種概念(species concept)」在鳥類分類學實踐中如何應用的一個絕佳教學案例。
1.5 全球分布範圍與台灣的記錄地位
大花鷚(狹義,即 Anthus richardi 全亞種)的全球分布範圍以東古北界(East Palaearctic)的開闊草地為核心繁殖地,越冬地則集中於印度次大陸、東南亞大陸及中國南方各地。繁殖地西起哈薩克斯坦東北部和西西伯利亞草原,向東延伸至外貝加爾地區(Transbaikalia)、鄂霍次克海沿岸、蒙古北部草原,以及中國東北部(黑龍江、吉林)和中國華南(揚子江以南地區);部分亞種的繁殖地也延伸至俄羅斯遠東的烏蘇里地區(Ussuriland)。越冬地主要為印度次大陸(從巴基斯坦至印度南部、斯里蘭卡)、孟加拉、緬甸、泰國、中南半島各國,以及中國華南沿海、海南島等地區,向南可達新加坡和婆羅洲北部。
台灣在大花鷚的分布記錄中,屬於其越冬範圍的東緣,被中華民國野鳥學會的《台灣鳥類名錄》(2023年版)定性為「冬候鳥(Winter Visitor)」,在實際出現的頻率與數量評估上則屬「不普遍(uncommon)」等級。這意味著大花鷚在台灣的冬季期間具有較規律的年度出現紀錄,並非偶發的迷鳥,但每個越冬季節的實際出現數量和分布地點均不穩定,因個體行為、氣候條件及覓食資源分布的年際變動而有所差異。台灣大花鷚的主要越冬記錄地點集中於西部及北部的低地平原農田、河濱草地和水邊荒地,以宜蘭平原、桃園大園一帶的農田、台南及嘉義的農業平原,以及台北盆地周邊的河濱公園草皮等地出現頻率較高。
第二章 演化簡史與多樣性、系統分類
大花鷚的演化歷史,是鷚屬在古北界草原生態系中輻射擴散的縮影,也是鳥類分類學在「物種界定(species delimitation)」這一核心議題上長達一個世紀爭論的活生生案例。理解這段歷史,不僅有助於認識大花鷚本身的自然史,更能幫助我們理解鳥類系統學如何在形態、分子和生態數據的交匯中不斷修正對自然界的認識。
2.1 鷚屬Anthus的演化起源:東亞起源假說與中新世輻射擴散
鷚屬(Anthus)是鶺鴒科中多樣性最高的屬,也是全球分布最廣的雀形目屬之一,其演化起源問題在鳥類學界長期以來是一個尚未完全解決的研究課題。根據現有的分子系統學分析,鷚屬的最近共同祖先(MRCA)起源時間估計在中新世(Miocene)晚期至上新世(Pliocene)初期之間,即距今約600至800萬年前,這一時期正值東亞及非洲草原生態系大規模擴張的地質時代。
分子親緣關係樹(molecular phylogeny)的研究結果,對鷚屬演化歷史提供了幾個重要線索。首先,非洲鷚類群的多樣性最高(非洲大陸擁有超過15種鷚屬物種),這一現象支持非洲可能是鷚屬早期多樣化的重要中心之一的假說;然而,部分基於分子定年的研究提出,鷚屬的起源中心可能在亞洲東部,其後分別向非洲和歐洲輻射,這一「東亞起源假說」目前在學界獲得一定程度的支持,但尚未形成完全的共識。其次,鷚屬並非一個單源(monophyletic)的嚴格自然類群——在某些分子系統樹中,鶺鴒屬(Motacilla)和長爪鷚屬(Macronyx)的部分物種與鷚屬成員混雜在一起,顯示鷚屬的傳統形態定義(以褐色縱斑羽色和地棲習性為主要特徵)可能是一種「功能性收斂演化(functional convergence)」的結果,而非嚴格的單一共同起源。這一發現提示,鶺鴒科各屬之間的系統關係遠比表面的形態相似性所暗示的更為複雜。
大花鷚在鷚屬的系統位置,根據現有的分子系統學資料,與布萊氏鷚(Anthus godlewskii)互為姐妹群(sister group),兩者共同構成一個更大的「大型草原鷚類」支系,再與稻田鷚(Anthus rufulus)及澳大拉西亞鷚(Anthus novaeseelandiae)等形成親緣較近的廣義近緣種群。這一系統位置解釋了大花鷚與布萊氏鷚在外觀上幾乎令人無法區分的形態相似性:兩者不僅在演化上是彼此最近的親戚,其分化時間也相對較晚,估計在更新世期間,當時的古北界草原生態系因冰期—間冰期的氣候波動而反覆擴張和收縮,造成鷚屬族群的反覆隔離與連通,可能促成了多個形態高度相似的姐妹種的分化。
2.2 鶺鴒科的系統位置與鶺鴒、長爪鷚的親緣關係
鶺鴒科(Motacillidae)在雀形目的系統位置,在分子系統學時代之前長期存在爭議。傳統的形態學分類曾將鶺鴒科置於「鶲形亞目(Oscines)」中一個相對孤立的位置,或與燕科(Hirundinidae)及其他在開闊環境活動的鳥類相鄰。現代分子系統學研究則將鶺鴒科明確置入「鴉雀總科(Passeroidea)」,與燕雀科(Fringillidae)、文鳥科(Estrildidae)、梅花雀科等構成一個較大的單源支系,反映了這些科之間在演化上的共同起源。
科內的系統關係方面,鷚屬(Anthus)、鶺鴒屬(Motacilla)與長爪鷚屬(Macronyx)三者之間的確切關係,在不同研究中呈現出不同的拓撲結構,但多數研究同意鶺鴒屬與長爪鷚屬互為姐妹群,兩者再共同與鷚屬形成鶺鴒科的三大主要支系。長爪鷚屬(Macronyx)是一個純粹分布於非洲的屬,以體色鮮豔(黃色或橙色腹面)和後爪超長為特徵,長爪鷚屬的後爪特化程度甚至超過大花鷚,在鶺鴒科中最為顯著,被認為是對非洲高草地棲息步行生活的極致適應。
山鶺鴒屬(Dendronanthus)是鶺鴒科中分類地位最特殊的屬,全屬僅含一種山鶺鴒(Dendronanthus indicus),這是鶺鴒科中唯一高度樹棲性的物種,其系統位置在不同研究中或置於鷚屬基部,或獨立為姐妹群,其特殊的樹棲行為和獨特的水平搖尾習慣(而非鶺鴒科通常的垂直上下擺尾),使其在整個科的演化框架中成為一個有趣的演化特例。
2.3 大花鷚的亞種多樣性:五至七個亞種的地理分布與形態差異
大花鷚在現行主要分類系統(IOC、HBW)中,認定有五個主要亞種,而部分舊分類系統或名錄(如ADW動物多樣性網站)則列出多達七至九個亞種名稱,差異源於部分亞種之間的形態分化程度是否足以支持亞種地位的判斷分歧。以IOC認定的五亞種為基準,各亞種的分布及主要形態差異如下。
指名亞種 A. r. richardi(維埃約,1818年命名)分布於西西伯利亞草原南部至哈薩克斯坦東北部,向東至貝加爾湖西岸,越冬主要在西南亞(印度西北部、巴基斯坦),部分個體繼續向西偏移,是在歐洲形成新遷徙路線的主要族群來源,也是建立「理查先生」命名典故的模式亞種。指名亞種整體體色較深,上體褐色調濃郁,背部縱斑清晰。
A. r. dauricus(約翰森,1952年命名,模式產地:西外貝加爾的烏庫本)分布於外貝加爾地區至雅庫特,向東至鄂霍次克海沿岸,南至蒙古北部和中國東北黑龍江省西北部,越冬於南亞,是在台灣越冬的主要亞種之一。dauricus 亞種以上體縱斑相對較多(more streaking above)為鑑別特徵,在不同亞種的形態比較中,這一特點最為穩定可靠。
A. r. centralasiae(奇斯蒂亞科夫斯基,1928年命名)分布於哈薩克斯坦東部(薩亞薩盆地、天山東段)至蒙古西南部和中國北部(新疆、青海、甘肅),越冬於南亞。centralasiae 是五亞種中體型最大的,上體底色最淺,趨向沙土色(sand-coloured upperparts),在外觀上給人一種略為褪色的印象,這與其繁殖地的乾旱半荒漠草原環境的保護色需求相吻合。
A. r. ussuriensis(約翰森,1952年命名,模式產地:俄羅斯濱海地區波西耶特灣)分布於俄羅斯遠東阿穆爾河下游地區(lower Amur)、烏蘇里蘭(Ussuriland),向南至中國東部(四川至揚子江),可能亦包括朝鮮半島,越冬於東南亞,是台灣越冬記錄的主要亞種之一,也是東南亞地區各越冬地最常記錄的大花鷚亞種。
A. r. sinensis(波納巴特,1850年命名,依瓦厄里訂正模式產地為中國福州)分布於中國揚子江以南的華南地區,越冬同樣在東南亞。sinensis 亞種是五亞種中體型最小的,上體縱斑最不明顯(least streaking above),整體色調較淡,在形態上最接近稻田鷚(Anthus rufulus),這也是 sinensis 亞種在部分分類系統中被建議移至稻田鷚物種下的分類討論的形態基礎之一。
2.4 東亞地理分布格局:繁殖地(西伯利亞至蒙古)與越冬地(南亞、東南亞)
大花鷚繁殖地的分布格局,呈現出一個以西伯利亞南部草原為核心、向外輻射至東亞各開闊低地的廣大版圖。繁殖地的核心環境是開闊的溫帶草原(temperate grassland),包括貝加爾湖周邊的蒙古草原延伸帶、外貝加爾丘陵草地、西伯利亞低地草原,以及中亞哈薩克斯坦的半乾旱草原帶。在這些地區,大花鷚的繁殖密度通常在適當棲地中達到相對較高的水準,是當地夏季鳥類群落的常見成員之一。與其他許多在西伯利亞繁殖的候鳥不同,大花鷚的繁殖地並不深入北極凍原(tundra)地帶,而是集中於北緯40至55度之間的溫帶草地,這一緯度帶的夏季具有充足的昆蟲食物和較長的日照時間,非常適合地面巢鳥類的快速育雛。
越冬地的格局則呈現出「南亞型」和「東南亞型」兩個主要分支,大致沿著不同亞種的繁殖地分布而有所分化:指名亞種(richardi)和 centralasiae 亞種的越冬地偏向印度次大陸西北部(印度、巴基斯坦、孟加拉);dauricus 亞種的越冬地跨越南亞全境;ussuriensis 和 sinensis 亞種的越冬地則偏向東南亞大陸(緬甸、泰國、中南半島),與台灣在地理位置上最為接近。這一地理分布格局意味著,在台灣越冬的大花鷚個體,最可能的繁殖地來源是俄羅斯遠東的烏蘇里地區及中國東北至華東一帶,即 ussuriensis 亞種的主要繁殖範圍;華南地區繁殖的 sinensis 亞種個體同樣有可能出現在台灣的越冬記錄中,但 sinensis 亞種本身的繁殖地與台灣的地理距離較近,部分個體也可能直接就地越冬於中國大陸南方而不南下台灣。
在越冬地的棲地選擇方面,大花鷚明顯偏好開闊的低地農業景觀,包括水稻田(尤其是冬季乾燥休耕後的稻田環境)、甘蔗田收割後的殘株地、草地牧場、河濱沙洲和低矮草本植被的荒地。在印度,大花鷚是農村田野中最常見的冬候鳥之一,常見於農田間的灌溉水渠邊緣和乾燥的收割後農田中;在泰國,大花鷚多見於稻田地區和河邊草地;在台灣,以上述農業景觀和河濱草地為主要棲息地點,與在同域越冬的樹鷚(偏好灌木草叢邊緣)和赤喉鷚(偏好短草地和農田)在微棲地選擇上有一定的重疊,但大花鷚通常更傾向於選擇較開闊、植被較低矮的地面環境,與其直立站姿和地面搜尋覓食的行為策略相吻合。
2.5 與相似物種的分類學糾葛:布萊氏鷚與稻田鷚的種界爭議
大花鷚的分類學糾葛,在當代鳥類學中最具代表性的是其與布萊氏鷚(Anthus godlewskii,Blyth's Pipit)之間的種界(species boundary)問題。布萊氏鷚由英國鳥類學家愛德華·布萊思(Edward Blyth)於1867年依據蒙古標本描述命名,繁殖地集中於蒙古東部至中國北方的乾旱草原,越冬地主要在印度次大陸,生態習性和分布範圍與大花鷚高度重疊。
兩者在外觀上的相似程度,在鷚類辨識中是公認的難題之一。主要辨識差異如下:布萊氏鷚的嘴喙(bill)相對較短;腿(tarsus)和尾(tail)均比大花鷚略短;後爪(hindclaw)比大花鷚更彎曲(more curved)且相對更短;尾羽外側白色面積較少;上體縱斑較多。最重要的形態鑑別點,在於翼中覆羽(median wing coverts)的羽毛形態:在成年大花鷚個體中,翼中覆羽的深色中心朝羽毛末端方向呈「菱形末端尖銳化(pointed tip)」,即深色區域在接近羽尖時形成一個近尖形的收縮點;而在布萊氏鷚的成年個體中,翼中覆羽的深色中心末端呈「鈍端(blunt-ended)」,即深色區域在羽尖附近呈較平截的結束。這一翼覆羽形態差異,是目前文獻中最可靠的大花鷚—布萊氏鷚辨識依據,但需要在近距離、良好光線下仔細觀察翼面細節,對大多數野外觀察者而言仍具相當難度。聲音方面,布萊氏鷚的叫聲較為柔和、不那麼刺耳,而大花鷚的叫聲(詳見第六章)相對更具爆發性和穿透力,是兩者在野外辨識中最實用的區別特徵之一。
與稻田鷚(Anthus rufulus)的糾葛則主要發生在 sinensis 亞種的分類歸屬問題上。sinensis 亞種在體型上最接近稻田鷚,上體縱斑也較其他大花鷚亞種少,部分形態測量數據(如翼長、尾長)與稻田鷚的重疊範圍較大,加上兩者的越冬地高度重疊(東南亞稻田環境),使得在越冬地進行精確的物種鑑定時,sinensis 亞種的個體對觀察者形成相當的辨識挑戰。部分鳥類學者主張將 sinensis 亞種重新歸入稻田鷚,但IOC等主要分類系統目前仍將其保留在大花鷚種下,這一爭議的最終解決,有賴於更全面的分子遺傳學分析和繁殖生物學比較研究。
台灣在大花鷚的全球遷徙網絡中,處於東亞越冬範圍的東緣。從繁殖地的地理位置推算,前往台灣越冬的大花鷚個體,需要從俄羅斯遠東或中國東北的繁殖地出發,向南飛越中國大陸東部,穿越台灣海峽抵達台灣,或由中國東南沿海繼續向南偏移至台灣;另一可能是由中國華南繁殖的 sinensis 亞種個體直接越海抵台。台灣在東亞候鳥遷徙體系中是一個重要的「中繼站島嶼(stopover island)」和「最終越冬地(wintering destination)」的雙重角色,對許多從西伯利亞南下的候鳥而言,台灣是其南向遷徙的最後一段或倒數第二段旅程的落腳點。
台灣的大花鷚越冬族群由哪些亞種構成,目前缺乏系統性的亞種鑑定研究,從現有文獻推算,ussuriensis 亞種(繁殖地最接近台灣的亞種)應是主要族群來源,dauricus 亞種個體也可能出現,而 sinensis 亞種因繁殖地在中國華南,地理上最有可能出現在台灣,但其體型較小、縱斑較少的形態特點,可能使部分個體在野外被誤判為稻田鷚。對台灣越冬大花鷚個體進行系統性亞種鑑定的工作,至今仍屬空白,是未來台灣鳥類學研究的一個值得填補的知識缺口。
第三章 大花鷚外觀特徵
大花鷚的外觀,在台灣可見鷚屬物種中最具辨識優勢的一點,是其相對突出的體型。當大花鷚與台灣最常見的鷚類——樹鷚(Anthus hodgsoni)和赤喉鷚(Anthus cervinus)——出現在同一視野中時,體型的差距相當明顯,幾乎無需辨識其他細節即可立即察覺。然而,當大花鷚獨自出現時,體型優勢失去了參照,辨識便需要依賴更細緻的形態特徵——羽色斑紋、後爪形態、站立姿態,以及最重要的叫聲。
3.1 體型測量:體長、體重、翼展與「鷚屬最大型之一」的相對地位
大花鷚成鳥全長(自嘴尖至尾羽末端)為17至20公分,以平均值計算約18至19公分,是全球鷚屬中體型最大的物種之一,僅略遜於分布於南美洲的部分大型鷚種。體重測量值在25至36公克之間,中國大陸方面的精細測量記錄顯示平均體重約為32.17公克,這一數值在雀形目小型至中型鳥類中屬於中等偏重的水準,反映其相對緊實的體型結構。翼展(wingspan)約為29至33公分,翼長(自翼角至翼尖的折翼長)約87至100公釐,平均約92公釐(92毫米)。
其他形態測量數據方面,嘴峰長(culmen length,自嘴基至嘴尖)約17至19公釐,平均約18公釐,嘴喙粗壯程度在鷚屬中屬於較大的範疇;嘴寬度約4公釐,嘴厚度約4.3公釐。跗蹠長(tarsus length)約28至30公釐,平均約29.2公釐,這一數值相對體長而言較為修長,賦予大花鷚在地面站立時明顯「長腿」的視覺印象,是其直立站姿的關鍵解剖學基礎。尾長約69至74公釐,平均約71.6公釐,尾羽外側白色邊緣(outer tail feathers white)清晰可見,在起飛時閃現的白色尾緣,是許多鷚屬物種的共通特徵,也是辨識鷚類起飛方向的實用視覺線索。雌雄個體在體型和外觀上高度相似,是典型的雌雄同形物種,野外無法僅憑外觀區別性別。
3.2 羽色組成:褐、黃褐、黑條紋的開闊地保護色系統
大花鷚的整體羽色組成,是一個以「褐底黑紋」為核心的開闊地保護色系統,功能上服務於在枯草、乾燥泥地和農田殘株環境中的視覺融合。上體(upper parts,即頭頂、後頸、背部、腰部)底色為褐色至黃褐色(buffish-brown),上面覆蓋著清晰的深褐色至黑褐色縱斑(dark streaks),縱斑在頭頂和背部最為密集,腰部和尾上覆羽縱斑較少,顏色略轉偏黃褐。尾羽(rectrices)整體為褐色,最外側一對尾羽(outer tail feathers)帶有白色外緣,在閉合的尾部兩側呈現為兩條細白線,在起飛展尾時白色面積明顯增大。
腹面(underparts)方面,喉部(throat)淡白至淡黃褐色,無縱斑或縱斑極少;胸部(breast)色調偏黃褐色(buffy),具有明顯的深褐色縱斑,縱斑在胸部兩側最為密集,中央稍疏;腹部(belly)及脅部(flanks)底色趨向白色至淡黃白色,縱斑稀少或無,腹部中央通常近乎素白。眉紋(supercilium)為較淡的黃褐色至白色,從嘴基延伸至耳羽後方,是頭部最清晰的淡色紋路;眼線(eye stripe)為深褐色;顴紋(malar stripe)自嘴角向下延伸,為深褐色,是分隔喉部白色與胸部縱斑區的形態界線。腿部(tarsi)顏色為淡粉褐色至肉黃色,偏淡的腿色與許多鷚屬物種的暗色腿腳形成對比,在野外觀察時長腿的淡肉色也是辨識大花鷚的輔助視覺特徵。
嘴喙顏色為深褐色至黑褐色,下嘴(lower mandible)的基部帶有黃色至黃褐色(yellowish base),這一嘴基底色特徵在近距離辨識時具有一定的參考價值,但並非所有個體均清晰可辨。
3.3 頭部紋路:眉紋、眼紋、顴紋的辨識功能
大花鷚頭部的紋路格局,是鷚屬物種辨識的核心區域之一。頭部紋路系統由眉紋(supercilium)、眼線(eye stripe)、顴紋(malar stripe)三條主要紋路組成,彼此之間的寬度、顏色對比和延伸長度,構成了對辨識者而言最直觀的視覺線索組合。
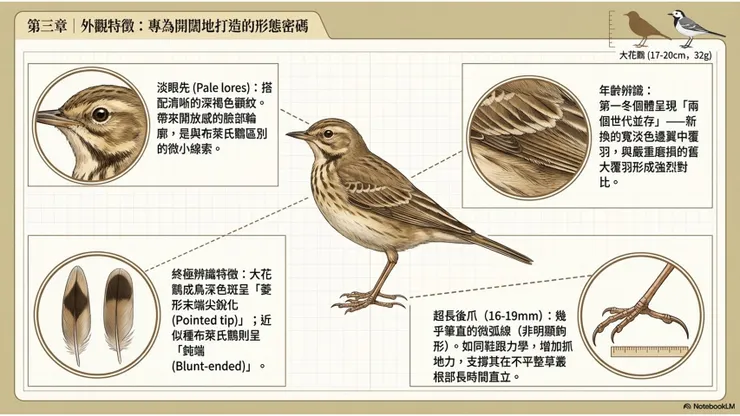
大花鷚的眉紋在整體鷚屬物種中屬於相對清晰且延伸較長的類型,從嘴基至眼先(lore)部分通常呈現為較寬的淡色帶,顏色為淡黃褐色至米白色,向後延伸至耳羽後緣;眼先(lores)部分的眉紋底色較淡(pale lores),這是大花鷚辨識的一個重要特徵——相對較淡的眼先部位,使頭部正面的臉部輪廓呈現出「開放感」,不同於部分鷚屬物種眼先偏深所造成的「封閉感」。深褐色眼線(從嘴基穿過眼部至耳羽的深色條紋)與淡色眉紋之間的對比,在大花鷚中屬於中等程度,不如赤喉鷚的眼紋那樣強烈,但仍清晰可辨。顴紋(malar stripe)在大花鷚中通常相當清晰,從嘴角沿著喉部邊緣向下延伸,深褐色的顴紋與喉部淡白色之間的對比,在近距離觀察時是一個可靠的辨識依據。
在與布萊氏鷚的頭部紋路比較中,大花鷚的眼先通常比布萊氏鷚更淡,呈現出更為明顯的「淡眼先(pale lores)」特徵,這一差異雖然細微,卻在仔細的野外觀察中具有一定的鑑別價值。
3.4 後爪(Hindclaw):超長後爪的形態特化與地面行走適應
大花鷚最重要的形態特徵之一,是其超長的後爪(hindclaw,即後趾爪)。後爪是第一趾(hallux,後趾)末端的角質化爪,在大花鷚中的長度達到16至19公釐,相對於整個後趾的長度(含趾骨和爪的總長度約25至28公釐),後爪的比例在鷚屬中屬於最長的類別。更重要的是,大花鷚後爪的形態特點不僅在長度上突出,還在彎曲程度(curvature)上與近似種形成明顯差異:大花鷚的後爪相對筆直(relatively straight),整體彎弧較小,幾乎近似一條微弧線,而非明顯的鉤形。相比之下,布萊氏鷚的後爪較短且彎曲程度更大,是最可靠的兩種區別特徵之一。
長而筆直的後爪的適應功能,首先體現在地面行走時的穩定支撐作用:超長後爪在行走時以爪尖輕觸地面、作為身體後方的穩定支點,類似於鞋跟的力學功能,使鳥在草地或泥地上行走時能夠保持更大的接觸面積,增加抓地力,減少在鬆軟或不平整地面上的滑倒風險。其次,長後爪也有助於在草叢根部的密集草莖環境中保持站立穩定性,使大花鷚能夠在不均勻的地面基質上長時間直立站立,執行其典型的「高挺站姿(upright posture)」行為。德文「Spornpieper(距鷚)」以這個超長後爪為命名核心,反映了這一形態特徵在歐洲早期博物學者眼中的顯著程度。
在野外辨識實踐中,後爪特徵的觀察需要相當近距離的視野(通常需要高倍望遠鏡或相機拍攝後放大),不如叫聲那樣在任何距離下均可立即利用。然而,在特別近距離的觀察條件下(例如河濱公園中的個體對人類警覺性相對降低時),長而筆直的後爪特徵確實能夠從外觀上直接確認大花鷚的身份。
3.5 翼覆羽菱形深色斑:區別大花鷚與布萊氏鷚的核心形態依據
如前文分類學章節所述,大花鷚成年個體翼中覆羽(median wing coverts)深色中心在羽尖方向呈現「尖銳菱形(pointed diamond shape)」的形態,是文獻中最被重視、最具可靠性的大花鷚—布萊氏鷚成年個體辨識特徵。這一特徵的觀察,需要在個體停棲、翼面折疊的狀態下,仔細比對翼上覆羽每根羽毛的深色中心形狀,具體而言即觀察每根翼中覆羽的深色區域,是在接近羽尖時逐漸收窄成一個相對尖細的端點(大花鷚),還是在羽尖附近以較寬的水平截面結束(布萊氏鷚)。
這一特徵的辨識挑戰在於:第一,翼覆羽的形態在不同光線條件下可能呈現出不同的視覺效果,羽毛表面的光線反射可能干擾對邊緣形狀的判斷;第二,翼覆羽的磨損程度(羽毛越磨損,尖端可能越不明顯)也會影響形態特徵的清晰度;第三,幼鳥(第一冬個體)的翼覆羽形態可能與成鳥有所不同,在年齡未確認的情況下,直接套用成鳥翼覆羽特徵進行辨識可能導致誤判。這些辨識挑戰,使得大花鷚—布萊氏鷚的精確辨識成為台灣賞鳥者在冬季田野中的重要技術挑戰,也是推動鷚類辨識技術進步的重要動力。
3.6 幼鳥、亞成鳥與成鳥的換羽差異及年齡辨識
大花鷚的年齡辨識,主要通過翼覆羽世代(covert generation)的對比來實現。在秋季抵達越冬地的個體中,「第一冬個體(first-winter)」即當年孵化、尚未完成第一次完整換羽的個體,其翼覆羽通常呈現出「兩個世代並存」的換羽對比(covert contrast):部分翼覆羽已完成換羽(新羽,通常為翼中覆羽),呈現出較寬的淡色羽緣(buff or white fringes),而另一部分仍保留幼鳥羽(juvenile coverts,通常為大覆羽 greater coverts),其淡色羽緣較舊、磨損程度較高,與已換新羽形成明顯的新舊對比。成鳥個體(second-year and older)在秋季換羽後,各翼覆羽世代趨於一致,淡色羽緣的磨損程度較為均勻,不呈現明顯的新舊對比。
幼鳥在孵化後最初的幾週,外觀上與成鳥相似,但整體色調更為均勻,斑紋相對較不清晰,嘴基黃色更為顯著,後爪雖然已相對較長,但尚未達到成鳥的充分發育長度。第一次完整換羽通常在到達越冬地後的冬季期間或次年春季在繁殖地到達後完成,換羽後外觀基本達到成鳥標準。在台灣越冬的大花鷚個體中,第一冬個體和成鳥個體通常混合出現,對有意深入研究台灣大花鷚族群年齡結構的賞鳥者和研究者而言,學習辨別兩種年齡類別是深化理解的重要一步。
第四章 大花鷚解剖與生理特徵
大花鷚的解剖與生理,在雀形目鶺鴒科的基本框架上,展現出與其長距離遷徙生活方式最直接相關的系統性特化——其中最重要的,是支撐長達數千公里遷徙飛行的脂肪積累機制、遷徙導航的神經生理基礎,以及昆蟲食性消化系統的構造。
4.1 視覺系統:開闊草地型鳥類的大視野全景視覺
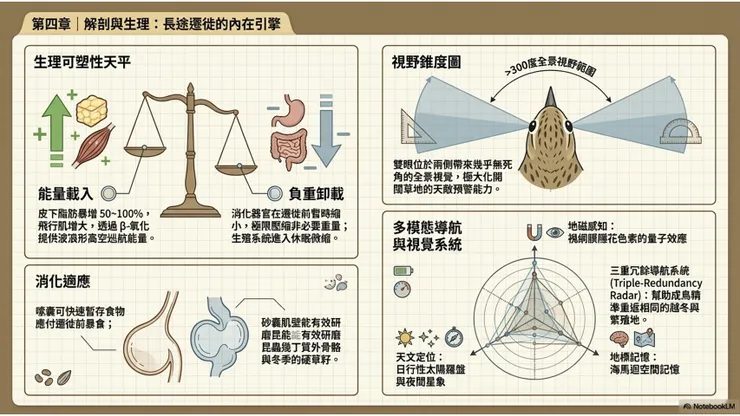
大花鷚棲息於開闊草地環境,在這種環境中,天敵(猛禽、猫科動物)的接近方向難以預測,唯有依靠寬廣的視野才能及早偵測威脅。鳥類的眼睛位置決定了其視野範圍:大多數鳥類(尤其是草地鳥類)的眼睛位於頭部兩側,提供接近360度的全景視野(panoramic vision),但兩眼正前方的雙眼重疊視野(binocular vision field)相對較窄,深度感知能力不如前向雙眼的猛禽。大花鷚的視覺系統符合這一草地鳥類的基本模式,其眼睛略大而圓,位於頭部側面,兩眼覆蓋的單眼視野(monocular field)各約150度以上,合計接近全方位覆蓋。
雀形目鳥類的視網膜含有四種錐狀細胞(相較人類的三種),能感知紫外線波段,這對在覓食時偵測昆蟲及其留下的尿跡(部分昆蟲的尿液在紫外線下具有反射特性)具有潛在的功能意義。視網膜中央凹(fovea)是視覺最敏銳的區域,大多數雀形目鳥類具有一個中央凹,而部分需要在開闊環境中快速偵測目標的鳥類具有兩個中央凹(bifoveal retina),其中一個針對正前方視野優化,另一個針對側向視野優化,但大花鷚是否具有雙中央凹結構,目前尚無針對性的解剖學研究資料。
4.2 骨骼系統與長距離遷徙的飛行力學
大花鷚作為長距離候鳥,其骨骼系統的基本特徵與所有鳥類相同:高度氣化(pneumatized)的骨骼,以輕量化的骨皮質在最小重量下提供足夠的機械強度。胸骨龍骨突(sternal keel)發育良好,支撐飛行主肌群(胸大肌和胸小肌)的附著,是鼓翼飛行動力的骨骼基礎。叉骨(furcula)在鼓翼週期中儲存和釋放彈性位能,提升長途飛行的能量效率。
大花鷚的翼形(wing shape)在鷚屬中屬於相對尖長型(pointed wing),翼長相對體重較大,翼展比(aspect ratio)偏高,這種翼型適合長距離高效率的持續飛行,在空氣動力學上能夠以較低的能耗維持穩定的長距離巡航,與需要在局部空間高機動飛行的短圓翼型(如大卷尾)形成對比。大花鷚的飛行模式為強而有力的波浪形鼓翼—滑翔交替節律,整體飛行特徵通常被描述為「強而起伏(strong and undulating)」,在田野上方做長距離移動時,波浪形的飛行軌跡是識別大花鷚起飛個體的一個行為特徵。
4.3 神經系統與地理空間記憶:遷徙導航的神經基礎
大花鷚進行每年往返長達數千公里的長距離遷徙,對神經系統提出了遠超日常棲居物種的認知要求。候鳥的遷徙導航系統被認為是多重感覺模態(sensory modality)的整合,包括地磁感知(magnetoreception)、天文定位(celestial navigation,包括太陽羅盤和星象定位)以及地理地標記憶(landmark memory)三個主要組成部分,這三個系統在不同的環境條件和遷徙階段中各自承擔不同比重的導航功能,並相互校正。
地磁感知是候鳥遷徙導航的基礎機制之一,目前研究支持的主要機制是眼睛中隱花色素(cryptochrome)光受體對地球磁場的量子效應感知(radical pair mechanism),使鳥類能夠感知地磁場的傾斜角(inclination),獲得一個類似「指南針」功能的方向感。遷徙前大花鷚的海馬迴(hippocampus)——即鳥類大腦中與空間記憶密切相關的腦區——可能通過繁殖地和越冬地的反覆到訪,建立並強化對關鍵地理位置的神經記憶,使成鳥個體能夠在多個遷徙季節後準確返回相同的繁殖地和越冬地。大花鷚幼鳥(第一次遷徙的個體)在初次南下時主要依賴遺傳編碼的方向程序(innate migratory direction program),逐漸在遷徙過程中積累地理記憶,轉向更精確的目標導向(goal-directed)導航模式。
4.4 消化系統:昆蟲食性的消化適應與偶爾取食種子
大花鷚以昆蟲和其他陸生無脊椎動物為主要食物,消化系統的構造反映了這一食性特點。砂囊(gizzard)的肌壁在大花鷚中屬於中等發育程度,能夠有效研磨昆蟲的幾丁質外骨骼,但不如以硬殼種子為主食的雀科(Fringillidae)鳥類那樣極端特化。大花鷚偶爾也取食草籽和其他小型種子,尤其在昆蟲食物較為稀少的越冬季節,種子可能在食物組成中占有更大比例,砂囊對種子的研磨能力提供了相應的消化支撐。
嗉囊(crop)能夠暫時儲存食物,允許個體在食物豐盛的地點快速積累,特別是在遷徙前的脂肪積累期,個體會增加取食頻率和每次取食量,嗉囊的食物暫存功能此時尤為重要。小腸是主要的營養吸收場所,腸道長度在雀形目中屬於中等,足以有效吸收昆蟲蛋白質、脂肪和碳水化合物。含氮廢物以尿酸的形式排出,尿酸的低水溶性使其能以半固態形式隨糞便排出,節省水分,這對在乾燥的草地環境中活動的大花鷚具有一定的滲透調節意義。
4.5 遷徙前的脂肪積累:皮下脂肪作為長途飛行能源的生理機制
大花鷚的長距離遷徙在能量上的核心挑戰,是如何在有限的體重中攜帶足夠的能量,以完成動輒數百至數千公里的連續飛行。候鳥解決這一問題的主要機制,是在遷徙前通過大幅增加取食頻率,在皮下組織(subcutaneous adipose tissue)和腹腔脂肪中積累大量脂質(lipid),作為長途飛行的主要燃料。
在啟程遷徙前,大花鷚個體的體重可能比非遷徙季節的正常體重增加50%至100%,以脂肪形式儲存的能量,在每公克脂肪能提供約9千卡熱量的基礎上,足以支撐一段長距離的連續飛行。在飛行過程中,脂肪通過β氧化(β-oxidation)途徑轉化為乙醯輔酶A,再進入三羧酸循環(Krebs cycle)產生ATP,為飛行肌提供持續的機械能輸出。在某些情況下,長途飛行的後期,當脂肪儲備接近耗盡時,飛行肌本身的蛋白質也可能被動員作為補充能量來源(蛋白質分解代謝),這一機制對器官功能有一定的消耗,需要在抵達中繼站或越冬地後盡快通過補食加以恢復。
此外,遷徙前後大花鷚的消化器官(尤其是砂囊、腸道)和飛行肌的相對質量會發生動態變化:在遷徙前積累脂肪時,飛行肌質量增加,而部分消化器官質量縮小(以減輕非必要的體重負擔);在抵達中繼站後,消化器官迅速恢復至正常大小,以支持快速補食。這種「器官大小可塑性(organ plasticity)」是長距離候鳥在遷徙—停棲—遷徙週期中調節身體資源配置的重要生理適應機制。
4.6 循環、排泄與生殖系統
大花鷚的心臟為四腔結構,靜息心跳頻率在每分鐘400至500次之間(根據體重推算),遷徙飛行時可進一步提升以應對飛行肌的高氧耗需求。鳥類的氣囊(air sac)呼吸系統確保肺部在吸氣和呼氣兩個相位中均能持續進行氣體交換,攝氧效率遠高於哺乳類的潮汐式呼吸,這對需要在數千公尺高空長時間持續飛行的大花鷚而言是重要的生理支撐。
大花鷚在台灣的越冬季節(10月至翌年4月)不進行繁殖,生殖系統在此期間處於非活躍狀態,雌鳥卵巢和雄鳥睪丸均縮小至最小體積,以減少維護非功能性性腺的能量消耗。隨著春季日照時間延長(光周期信號),個體在北返遷徙途中生殖系統逐漸啟動,抵達繁殖地時已達到可立即開始繁殖活動的生理準備狀態。
第五章 大花鷚生活史特徵
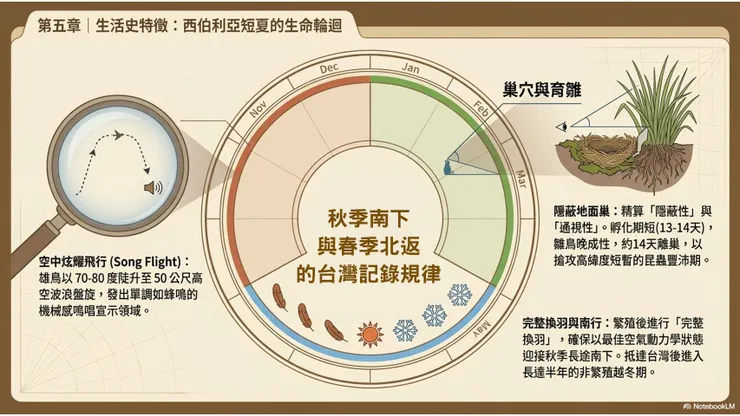
大花鷚的生活史,以遷徙週期為主軸,在西伯利亞繁殖地和南亞—東南亞越冬地之間,以近乎精確的年度節律往返。在台灣,觀察者所見的大花鷚,始終是處於「越冬」這個生活史階段的個體,其繁殖生活史的全貌,只能通過在遠方繁殖地進行的野外研究來加以揭示。
5.1 繁殖季節(5至7月)與繁殖地環境:西伯利亞草原的夏季生命
大花鷚的繁殖季節,集中在每年5月至7月之間,與西伯利亞夏季短暫的高緯度昆蟲豐盛期高度吻合。在貝加爾湖周邊、外貝加爾地區和蒙古北部草原,5月中下旬氣溫回升、積雪融化,草地植被開始萌發,隨之而來的是各類昆蟲(蚱蜢、甲蟲、蒼蠅、蛾類幼蟲等)族群的快速增長,為育雛提供了充足的高蛋白食物來源。大花鷚通常在5月底至6月初開始建立繁殖領域並啟動築巢,整個繁殖周期(從築巢至雛鳥獨立)在7月下旬至8月初大致完成,之後即陸續開始秋季南下遷徙準備。
大花鷚的繁殖地環境,是典型的「廣域開闊草原型(open steppe type)」,包括天然草原、草甸、牧草地和疏林草地的過渡帶,共同特點是植被高度低矮(通常低於30公分)、地面開闊,具有足夠的視野能讓親鳥及時發現接近巢位的天敵。在蒙古和外貝加爾地區,大花鷚的繁殖棲地與蒙古草原上的傳統遊牧放牧地高度重疊,放牧牲畜的踐踏雖然在一定程度上改變了草地的植被結構,但保持了低矮植被和開闊地面的物理環境特性,在某些情況下甚至對大花鷚的繁殖棲地質量具有中性或輕微正向的影響。
5.2 求偶展示:空中炫耀飛行(Song Flight)與單調蜂鳴鳴唱
大花鷚的求偶行為以空中炫耀飛行(song flight)為核心展示形式,這是鷚屬物種的共通繁殖行為特徵之一,在大花鷚中表現為雄鳥從地面起飛,以陡升的飛行軌跡上升至30至50公尺的高空,然後以拍翼—收翼交替的波浪狀飛行維持在高空盤旋,同時持續發出重複性的鳴唱;在展示完成後,雄鳥通常以雙翼收攏的俯衝姿態急速降落至地面。這種將高空曝光(增加可見度)與鳴唱(傳遞聲音信號)相結合的展示形式,使雄鳥能夠在開闊草地的廣大範圍內同時向雌鳥和競爭雄鳥傳遞繁殖狀態的信號。
大花鷚的鳴唱本身,被文獻描述為「一系列反覆的單調蜂鳴音(a repeated series of monotonous buzzy notes)」,與鴉科的複雜多元鳴聲或鶇科的悅耳旋律截然不同,大花鷚的繁殖期鳴唱音質相對單調,帶有一種規律性的機械感,類似某些昆蟲的振翅音。這種鳴唱的信息功能,主要是以持續時間和強度傳遞雄鳥的體力狀況和領域占有信號,而非以音樂複雜度吸引雌鳥。雌鳥在評估雄鳥的求偶展示後,以其自身的行為反應(接近、跟隨或逃離)傳遞接受或拒絕的信號;配對確立後,雄鳥的空中炫耀飛行頻率通常有所下降,但在繁殖季節仍會持續進行以維繫領域宣示的功能。
5.3 巢位選擇:草叢根部的隱蔽地面巢
大花鷚採典型的地面築巢(ground nesting)策略,與其草地棲息習性高度匹配。巢位通常建立在一叢較高的草或灌木根部,利用草叢的基部空間作為掩護,從上方或前方難以直接看見巢的位置,保護色效果良好。這種隱蔽型的巢位選擇,是地面巢鳥類應對草地環境中天敵壓力(包括蛇類、小型哺乳動物和大型昆蟲食性鳥類)的主要防衛策略,以視覺隱蔽代替主動防衛,儘量降低巢位被發現的機率。
巢址選擇時,大花鷚成鳥對周圍地面植被的高度和密度有一定的偏好:過於稀疏的地面植被無法提供足夠的遮蔽;過於茂密和高大的植被則妨礙親鳥的視野和進出路線,不利於快速偵測天敵。草叢根部的空隙提供了「隱蔽性(concealment)」和「通視性(visibility)」之間的恰當平衡,是大花鷚巢位選擇的核心邏輯。在台灣,並無大花鷚繁殖的記錄,其地面築巢的相關觀察只能依賴其在繁殖地(西伯利亞、蒙古)所進行的研究資料。
5.4 產卵、孵化期(13至14天)與雙親育雛分工
大花鷚每窩通常產卵3至5枚,以4枚最為常見。卵的外觀為具有細緻斑紋的保護色型,底色為灰白色至淡褐色,上面覆蓋著深褐色至灰黑色的細密斑點和斑紋,斑紋往往在鈍端(較寬的一端)更為密集。卵的長徑約22至24公釐,短徑約16至17公釐,重量約3至4公克。這種保護色卵的視覺效果,與草地地面的枯葉和泥土背景高度融合,在親鳥離巢時使巢中的卵極難被陸地天敵在視覺上發現。
孵化期(incubation period)約13至14天,在鷚屬物種中屬於較短的範疇,反映了高緯度繁殖地夏季短暫、雛鳥必須盡快成長的時間壓力。孵化工作以雌鳥為主,雌鳥具有發育的孵卵斑(brood patch),能夠以腹部皮膚直接接觸卵並加熱,維持孵卵所需的恆定卵溫。雄鳥在孵化期的主要貢獻包括:在雌鳥孵卵時擔任巢址周邊的警戒哨兵;在雌鳥需要外出覓食時,偶爾替代雌鳥在巢上短暫保溫;以及積極驅趕接近巢位的天敵。雌雄鳥在育雛期均承擔覓食和餵食雛鳥的工作,兩者的外出覓食頻率隨雛鳥生長和食物需求量增加而持續上升,育雛高峰期的親鳥覓食頻率可達每小時數次。
5.5 雛鳥發育、離巢(約14天)與幼鳥獨立過程
大花鷚雛鳥為晚成性(altricial),孵化時幾乎無羽,眼睛閉合,完全依賴親鳥的保溫和餵食。孵化後第一週生長最為迅速,體重快速增加,羽囊(feather follicles)逐漸顯現,針羽(pin feathers)開始萌出;第二週,正羽(contour feathers)加速發育,雛鳥體溫調節能力逐漸建立,親鳥的護雛(brooding)時間相應縮短。雛鳥在孵化後約12至14天具備初步飛行能力後離巢,但此時飛行能力尚不完善,離巢後仍繼續在巢址附近的草叢中活動,依賴親鳥持續餵食。
離巢後的幼鳥,在親鳥的引導下逐漸學習地面搜尋昆蟲的覓食技能,這一學習期約持續2至3週,幼鳥的取食獨立性逐漸提升,親鳥的餵食頻率隨之降低,直至幼鳥完全自食其力。通常在離巢後約4週,幼鳥已基本達到食物自給的能力,此後親鳥的投資進一步減少,親子關係逐漸解除。若第一窩繁殖順利,西伯利亞繁殖地的大花鷚有時會在6月下旬至7月初啟動第二窩繁殖,但高緯度繁殖地的短暫夏季使第二窩繁殖的時間餘裕相當有限,部分個體或許更傾向於把握秋季食物豐盛期進行充分的脂肪積累,而非追加第二窩。
5.6 換羽週期、成鳥期與野外壽命估計
大花鷚的換羽週期,以繁殖季節結束後的「繁殖後完整換羽(post-breeding complete molt)」為主要換羽事件,通常在繁殖地完成或在遷徙途中的中繼站進行。完整換羽使所有飛羽、尾羽和體羽更新,確保個體以最佳的羽毛狀態啟程進行秋季遷徙。幼鳥(當年孵化個體)在離巢後進行「幼後換羽(post-juvenile partial molt)」,更換大部分體羽,但通常保留部分飛羽和翼覆羽,形成前述的「翼覆羽世代對比」。大花鷚在約一歲至一歲半時達到性成熟,開始首次繁殖。野外壽命估計在4至7年之間,但確切數據因缺乏系統性的繫放長期追蹤研究而難以精確確認。主要死亡壓力包括遷徙途中的惡劣天氣(暴風雪、強烈側風)、能源耗盡、在繁殖地和越冬地的天敵捕食,以及棲息地質量下降造成的食物可獲性降低。
第六章 大花鷚行為生態
大花鷚在田野中的行為,有一種低調而自信的質感。牠不像大卷尾那樣主動宣示存在,也不像台灣藍鵲那樣以群體行動形成視覺上的衝擊力;牠只是靜靜地站在那裡,筆直,高挑,以一種幾乎刻意的警覺姿態環視周圍,然後走兩步,停一下,再走兩步——在台灣冬季田野的安靜角落裡,如果你知道如何尋找,就有機會看見這個來自遙遠西伯利亞的旅人在此短暫停棲的身影。
6.1 棲息地選擇:繁殖地與越冬地的開闊低地偏好
大花鷚在繁殖地和越冬地的棲地選擇,均以低矮植被的開闊低地為核心,這一高度一致的棲地偏好貫穿了其全年生活史的各個階段。在繁殖地,開闊草地提供了築巢所需的地面植被掩護和充足的昆蟲食物;在越冬地,低矮農業植被(稻田、旱田殘株)和草地提供了同樣的地面昆蟲食物來源,以及足夠的視野供個體警覺天敵。
在台灣的越冬棲地選擇中,大花鷚主要出現於:(1) 西部及北部平原的水稻田,尤其是冬季乾燥休耕後的稻田,地面出露、植被矮短,適合地面步行覓食;(2) 河濱公園和堤防草皮,以短草坪環境為主,常與赤喉鷚、東方黃鶺鴒等同域越冬鳥類共享棲地;(3) 農業灌溉渠道和田埂邊緣的草地;(4) 沿海低地草地和廢耕農地。值得注意的是,大花鷚在台灣的分布雖遍及全島低地,但並非在所有看似合適的農田草地都能穩定記錄到,可能反映了其在台灣越冬個體數量本身的有限,以及個體在越冬地點選擇上的一定隨機性。
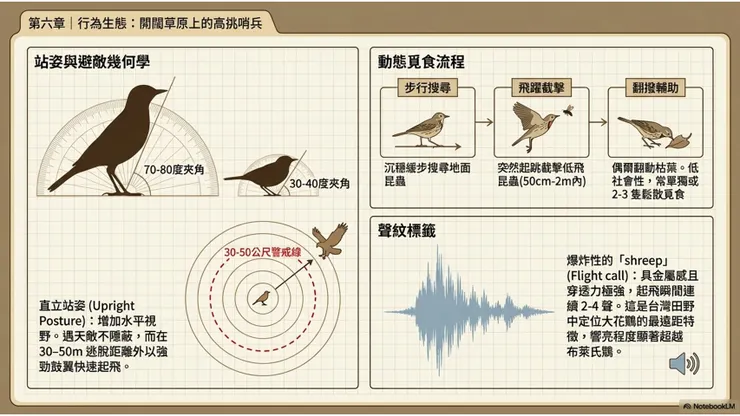
6.2 直立站姿(Upright Posture):大花鷚最具辨識性的行為特徵
在台灣田野觀鳥的經驗中,能讓有經驗的觀鳥者第一時間辨識大花鷚的,往往不是其羽色細節,而是一個動作特徵——那種近乎誇張的直立站姿(upright posture)。當大花鷚停在地面上時,其身體軸線幾乎與地面呈70至80度的夾角,頸部微微伸直,頭部高舉,給人一種「鶴立雞群」式的頎長感。相比之下,同樣在草地活動的赤喉鷚、樹鷚和雲雀(Alauda arvensis)等鳥類的站立姿態相對更為水平傾斜,沒有大花鷚那種近乎直立的挺立感。
這種直立站姿的行為生態學意義,首先是警覺功能:在開闊草地環境中,直立姿態使鳥類的眼部高度更高,能夠獲得更遠的水平視野,更早發現遠距離的潛在天敵或同種競爭者。其次,直立站姿也可能具有社交信號功能,在個體之間的短距離對峙情境中,更高的站立姿態可以增加自身的視覺體積感,有效傳遞「我比你更大」的直觀信號。大花鷚在受到干擾時,典型的反應是先靜止直立(freeze response),以最高的身體姿態掃視威脅來源,判斷威脅等級後再決定是繼續停留(若威脅程度低)還是起飛逃離(若威脅接近警戒距離)。
6.3 地面行走與覓食策略:步行搜尋、短暫飛躍截擊與地面翻撥
大花鷚的主要覓食動作是地面步行搜尋(ground walking search),即以緩慢、穩定的步伐在地面上前進,以眼睛持續搜尋地面昆蟲,發現目標後以嘴喙迅速啄取。這種步行覓食模式在鷚屬物種中是共通的基本覓食策略,適合在昆蟲密度均勻分布的草地環境中系統性地搜索食物,能量效率介於「守株待蟲(sit-and-wait)」和「高速追逐(high-speed pursuit)」兩種極端策略之間的中間層次。
在昆蟲目標飛起或進行短距離移動時,大花鷚會從地面起跳,以短暫飛躍動作(short aerial lunge)截擊飛行中的昆蟲,然後立即降落回地面繼續步行搜尋,這個空中截擊動作的飛行距離通常在50公分至2公尺以內。部分文獻也描述了大花鷚偶爾在草叢邊緣翻撥枯葉(leaf-turning)以搜尋藏匿其下昆蟲的行為,雖然此行為在大花鷚中的發生頻率相對低於林地昆蟲食性鳥類(後者高度依賴翻葉覓食),但在食物較為分散的越冬地草地環境中,翻撥地表覆蓋物也是補充覓食的輔助策略之一。
在台灣觀察到的大花鷚個體,覓食時通常獨自活動,以緩步行走為主,偶爾停下直立警覺,再繼續前進,整體覓食行為節奏沉穩而有規律。在食物豐盛的地點(如翻耕農田),有時可見幾隻大花鷚在相對接近的距離內各自覓食,但個體之間維持一定的距離,不顯示明顯的協同覓食傾向。
6.4 社群行為:越冬期的鬆散小群與遷徙期的聚群動態
大花鷚在越冬地的社群結構,以鬆散的個體獨立活動為基本模式,有別於部分高度群居的雀科鳥類。在台灣越冬的大花鷚,通常以單獨個體或2至3隻的鬆散小群出現,偶爾在同一農田範圍內可見數隻個體分散覓食,彼此之間維持一定距離,不形成緊密的社會性群體。這種低社會性的越冬行為,與其在廣大草地上分散分布的覓食需求相吻合:昆蟲食物在草地上的分布相對分散,不需要群體合作才能有效獲取,群體聚集反而可能增加食物競爭。
在遷徙途中,大花鷚的社群性略有提升,遷徙中的個體有時會與其他同種個體或近親物種(如布萊氏鷚、赤喉鷚)形成較大的鬆散遷徙群,共同利用遷徙中繼站的食物資源。這種暫時性的遷徙聚群,在停棲休息時最為明顯——多隻個體可能同時停棲在同一農田或草地,形成視覺上相對密集的分布,但個體之間的互動仍以避讓(agonistic spacing)為主,缺乏明顯的協同行為。大花鷚在遷徙期的記錄,在台灣主要集中在秋季(9月中旬至11月)和春季(3月至4月),特別是在台灣西部海岸線和農田地帶,這些地點是東亞鳥類沿海遷徙走廊的重要組成部分。
6.5 叫聲生態:爆炸性「shreep」鳴叫的功能、傳播距離與辨識價值
大花鷚的叫聲在台灣越冬地的野外辨識中,是最具決定性意義的辨識特徵之一,其重要性甚至超越所有形態特徵。大花鷚最常發出的叫聲,被英文文獻描述為「爆炸性的 'shreep'」(explosive 'shreep'),這個單音節叫聲音量大、頻率高、穿透力強,音質帶有一種硬而尖銳的金屬感,與麻雀的「啾」聲在音質上有一定的相似性(文獻描述「與家麻雀的叫聲有幾分相似 somewhat similar to the chirp of a house sparrow」),但音量顯著更大、更具爆發力。
在台灣田野中,大花鷚的「shreep」叫聲通常在個體受到驚嚇起飛時最為頻繁,一隻被驚動的大花鷚在起飛的瞬間,往往會連續發出2至4聲響亮的「shreep、shreep、shreep」,這種起飛叫聲(flight call)在數十公尺外均清晰可辨,即便在觀察者尚未在視野中確認個體時,叫聲已先行宣告了大花鷚的存在。有經驗的賞鳥者,往往是先聽到這個叫聲,才去尋找起飛個體的飛行方向,進而目視確認。
與布萊氏鷚的叫聲比較,大花鷚的「shreep」相對更為響亮、更具爆發性和穿透力,而布萊氏鷚的叫聲(文獻描述為相對更柔和 'quieter and less harsh')在音量和尖銳度上均略遜於大花鷚,是兩者在聲音特徵上最可靠的辨識差異。稻田鷚的叫聲則更短促,聲音品質更為柔弱,與大花鷚的爆炸性叫聲差異更大。掌握大花鷚叫聲的識別,是台灣賞鳥者在冬季田野中高效定位大花鷚的最重要技能。
6.6 天敵迴避:開闊環境下的警覺策略與快速起飛逃脫行為
在開闊的農田草地環境中,大花鷚的天敵壓力主要來自日行性猛禽,其中紅隼(Falco tinnunculus)、東方澤鵟(Circus spilonotus)和遊隼(Falco peregrinus)是台灣冬季農田中最常見的小型鳥類天敵。面對這些空中威脅,大花鷚的防禦策略以「早期偵測+快速起飛逃脫」為核心,而非依賴靜止隱蔽(freeze-and-hide),這與其所棲息的開闊無遮蔽環境高度相符——在開闊地面,靜止不動既不能有效遮蔽,也無法在最後一刻快速移動至掩護處,因此以距離和速度為基礎的飛行逃脫是最實際的防禦選擇。
大花鷚的直立站姿(前述),使其在地面覓食時能夠保持對周圍廣大空間的持續視覺監控,通常在天敵靠近到約50至80公尺的警戒距離(alert distance)時即停止覓食,轉入警覺模式(freeze posture);當天敵繼續靠近至逃脫距離(flight initiation distance,通常約30至50公尺)時,大花鷚迅速起飛,以爆炸性的強勁鼓翼迅速爬升至數公尺的飛行高度,然後轉向遠離天敵的方向快速離去,同時發出一串「shreep」叫聲。起飛後的逃脫飛行通常距離較長(50至200公尺),大花鷚不會立即降落在原地附近,而是飛越較大距離後降落在另一塊農田或草地,以確保與天敵之間維持足夠的安全間距。
第七章 大花鷚遷徙生態學
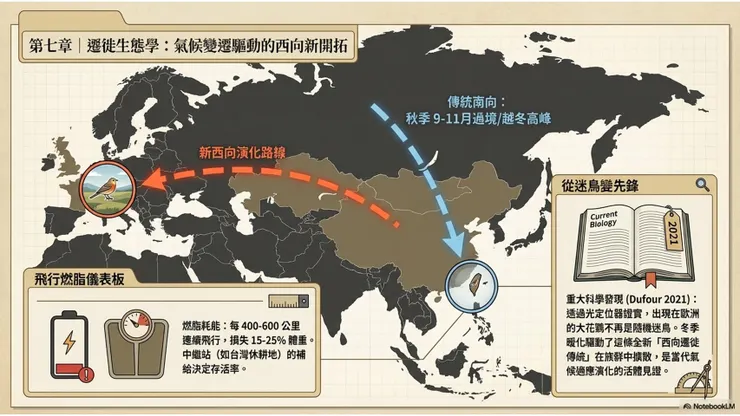
大花鷚的遷徙,是其生命故事中最引人入勝、在科學上最具前沿研究價值的面向。每年秋天,來自西伯利亞草原的大花鷚個體匯聚成一道無形的候鳥之流,向南飛越中亞、中國大陸,抵達印度次大陸和東南亞的越冬地;春天,這條流向逆轉,個體重返數千公里外的繁殖地。然而近年的科學研究揭示,除了這條傳統的南北向主路線,部分大花鷚個體正在開拓一條全新的、方向幾乎截然相反的遷徙路線——向西,跨越整個歐亞大陸,抵達西歐越冬。這個發現,在當代鳥類遷徙科學的框架中,是理解遷徙行為如何演化、以及氣候變遷如何重塑動物行為的一個里程碑式案例。
7.1 遷徙路線:南向主幹道(東南亞度冬)與西向新路線(歐洲)的雙軌系統
大花鷚的傳統主遷徙路線,以「南向(southward)」為基本方向,從西伯利亞和蒙古的繁殖地出發,秋季沿著東亞季風走廊向南飛行,穿越中國大陸(通過華北平原、長江流域、華南沿海等主要停棲中繼站),最終抵達印度次大陸、東南亞大陸(緬甸、泰國、中南半島)及中國南方各地的越冬地,部分個體繼續向南延伸至新加坡和婆羅洲北部。台灣位於這條南向主路線的東緣,接收來自中國東北和華東方向南下的部分個體。
然而,大約在20世紀後半葉,歐洲的觀察者開始注意到,每逢秋季,在英國、法國南部、伊比利亞半島等西歐地區,有越來越多的大花鷚個體出現在當地的草地和農田。最初,這些個體被視為「迷鳥(vagrant)」——即偶爾偏離正常遷徙方向的「迷途」個體,這在候鳥遷徙記錄中是常見現象。然而隨著西歐各地大花鷚記錄數量的逐年增加,科學家開始懷疑這是否僅僅是迷鳥數量的隨機增長,還是背後有更系統性的原因。
7.2 2021年《當代生物學》研究:氣候變遷驅動的西向遷徙新路線演化
2021年10月,法國博物館博物學家Dufour等人在《當代生物學》(Current Biology)期刊發表了一篇具有里程碑意義的研究論文,題為「亞洲雀形目鳥類的新西向遷徙路線(A new westward migration route in an Asian passerine bird)」。這篇論文通過一系列嚴謹的野外追蹤和標記研究,首次科學確認了大花鷚在西歐出現並非單純的迷鳥現象,而是代表了一個真實建立的新遷徙路線(new migration route)——一個正在當代時間尺度上發生的遷徙行為演化事件。
研究者首先對在法國南部越冬的大花鷚個體進行了繫放標記,記錄到同一批個體在連續多個冬季返回法國南部越冬,這直接反駁了「每年出現的都是不同的隨機迷鳥」的假說——如果是隨機迷鳥,不應有個體重複出現在同一地點的模式。其次,研究者在部分個體身上安裝了光定位器(geolocators),追蹤這些個體的全年移動路徑,結果顯示:在法國南部越冬的大花鷚個體,其春季北返的路徑是向東穿越中歐,回到中亞(哈薩克斯坦東部至蒙古邊境地帶)的繁殖地,整個遷徙旅程的距離長達約6000公里,且方向是罕見的「緯向西向(longitudinal westward route)」,即在緯度變化不大的情況下,沿東西方向橫越整個歐亞大陸,這在已知的亞洲遷徙鳥類中屬於極不尋常的遷徙路線類型。
這一新遷徙路線的形成,被研究者與氣候變遷(climate change)緊密關聯。大花鷚繁殖地西緣(哈薩克斯坦東部至西西伯利亞草原)的氣候在過去幾十年間發生了顯著變化,冬季溫度上升使西歐越冬地的環境條件更加適宜,可能降低了向西遷徙的能量代價和存活風險;同時,氣候驅動的食物可獲性變化,可能改變了特定繁殖地族群的遷徙方向偏好,使部分個體偏離傳統南向路線,向西偏轉進入歐洲。一旦這些「先鋒個體(pioneer individuals)」成功在西歐越冬並安全返回繁殖地完成繁殖,其遷徙行為(無論是遺傳編碼的還是社會學習傳遞的)就有機會傳遞給後代,形成一個新的遷徙傳統(migratory tradition)在族群中擴展。
這項研究的科學意義,遠超大花鷚本身的物種研究:它首次在當代時間尺度上,通過追蹤數據直接記錄到了一個新遷徙路線正在鳥類族群中建立的過程,這在動物行為演化研究中是前所未有的直接觀察機會。《當代生物學》配套的評論文章(Lees & Gilroy,2021)將此發現定性為「當迷鳥變成先鋒(When vagrants become pioneers)」,精準地概括了從隨機迷鳥到有目的遷徙先行者的關鍵概念轉變。
7.3 遷徙時序:秋季南下(9至11月)與春季北返的台灣記錄規律
在台灣,大花鷚的季節性出現規律,與其在東亞遷徙走廊中的時序相吻合。秋季南向遷徙的台灣過境和越冬期,通常在9月中旬至11月初之間,此時來自西伯利亞和蒙古的大花鷚個體陸續出現在台灣北部和西部的低地農田;在台灣全島範圍內,10月至11月是秋季記錄頻率最高的時段,與東北季風開始盛行的時間高度吻合,強勁的東北風幫助候鳥的南向遷徙。部分個體在台灣越冬,留至次年3至4月的春季北返期;春季台灣的大花鷚記錄通常少於秋季,可能反映了個體在春季北返時的飛行速度更快(繁殖時間壓力驅動的快速春季遷徙),在台灣停留時間更短,單日被觀察到的機率下降。
冬季(12月至翌年2月)的台灣大花鷚記錄,雖不穩定,但在宜蘭平原農田、台南西部沿海農地及彰化農業平原等地,有年度越冬個體的相對穩定記錄,顯示台灣確實有少量個體選擇在此越冬而非繼續南下至更南方的越冬地。台灣作為越冬地的吸引力,可能與台灣冬季平均氣溫相對較高(北台灣低地冬季通常在攝氏12至20度之間)、農業昆蟲食物在冬季仍有一定可獲性有關。
7.4 遷徙中的脂肪燃燒、體重管理與中繼站選擇策略
大花鷚在遷徙過程中的能量管理,是其遷徙成功的核心生理挑戰。在啟程前積累的脂肪儲備,在長距離飛行段落中以高效率燃燒,為飛行肌持續提供能量。一隻體重32公克的大花鷚,在遷徙前積累的脂肪可能使體重增加至50公克以上;在一段400至600公里的連續飛行後,燃燒的脂肪可能使體重下降5至10公克,即損失約15至25%的總體重。抵達中繼站後,個體需要盡快補充食物以恢復脂肪儲備,中繼站的食物可獲性因此成為決定個體遷徙速度和整體遷徙成功率的關鍵因素之一。
中繼站的選擇策略,在大花鷚中遵循「食物最大化(food maximization)」的基本邏輯:個體傾向於選擇昆蟲食物豐盛、天敵壓力低的開闊低地農田或草地作為中繼站停棲地。在台灣,大卷尾越冬記錄較多的幾個點(宜蘭平原農田、西部沿海農地),也常常是大花鷚中繼站停棲的熱點,顯示這些地點在食物可獲性和環境條件上,對多種候鳥均具有吸引力。在中繼站的停留時間,從一天至數天不等,取決於個體的脂肪儲量、食物可獲性,以及天氣條件(強順風天氣下個體傾向快速啟程,逆風或暴風天氣時個體傾向延長停留等待)。
7.5 遷徙導航機制:地磁感知、星象定位與地標記憶的多重系統
大花鷚的遷徙導航系統,綜合運用了鳥類已知的多種導航機制,形成一個具有冗餘(redundancy)的多模態導航體系。地磁感知(magnetoreception)提供了基本的方向感知,使鳥類即便在陰雲密布、無法見到太陽或星星的夜間飛行條件下,仍能維持大體正確的遷徙方向。太陽羅盤(sun compass),即利用太陽在天空中的方位和仰角推算方向,在日間遷徙時提供精確的方向校正。星象定位(star navigation),即利用夜空中恆星的圖案(尤其是北極星周邊的星群)確認正北方向,對許多在夜間遷徙的候鳥(大花鷚為日行性遷徙者,夜間遷徙的比例較低,但仍可能在部分條件下進行夜間飛行)具有重要的補充功能。
地理地標記憶(landmark memory)在大花鷚成鳥的遷徙中可能扮演重要的細化導航角色:在完成第一個繁殖季節後,成鳥已積累了往返遷徙路線上關鍵地理特徵的記憶(如海岸線走向、主要河流、山脈輪廓),這些記憶有助於個體在熟悉的路線區段內以精確的地標導航代替純粹的地磁或天文方向感知,從而實現更精確的中繼站再訪。幼鳥(初次遷徙個體)主要依賴遺傳編碼的方向偏好(initial bearing preference),在尚未積累地理記憶的情況下,以「遷徙向量(migration vector,即方向加距離的組合)」指引初次遷徙旅程,雖然成功率低於成鳥,但足以使多數個體抵達越冬地的大致範圍。
第八章 大花鷚保育生態學
大花鷚的保育生態學,在台灣的語境下,更多是一個關於「如何確保這個不普遍的冬候鳥在台灣持續有機會被觀察和研究」的問題,而非緊迫的瀕危物種拯救課題。然而在其繁殖地(西伯利亞和蒙古草原),以及越冬地(印度次大陸和東南亞農業景觀)的環境品質,以及氣候變遷對其遷徙生態的長期影響,構成了大花鷚保育問題的真正核心所在。
8.1 生態服務功能:開闊草地昆蟲調控與食物網角色
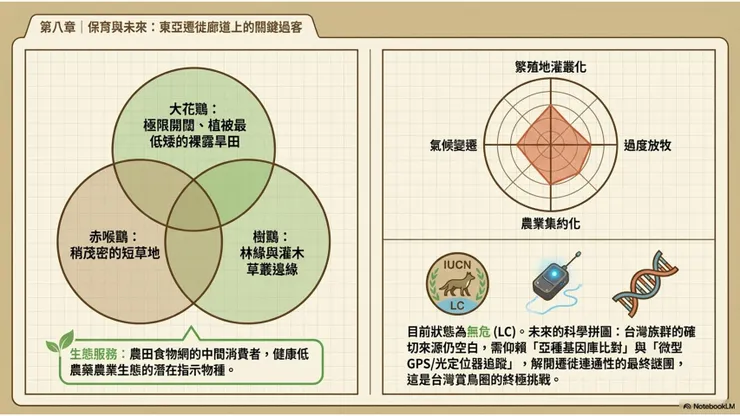
大花鷚在其越冬地的生態服務功能,以昆蟲消耗作為最主要的貢獻。在印度次大陸和東南亞農業景觀中,大花鷚的越冬密度有時相當可觀,其對農業昆蟲的消耗,尤其是對越冬農田中各類草地昆蟲幼蟲和成蟲的取食,對局部農業害蟲的自然防治具有一定的正向貢獻。雖然與常年在農田活動的大卷尾(在台灣扮演更為穩定和顯著的農業益鳥角色)相比,大花鷚作為冬候鳥的昆蟲消耗貢獻在時間上受到越冬季節的限制,但在昆蟲消耗的時間段(秋冬季節)和棲地類型(開闊農田地面)上,大花鷚的貢獻具有一定的補充意義。
在食物網的位置方面,大花鷚在台灣越冬期間是農田食物鏈的中間消費者:捕食各類地面昆蟲(初級消費者層次),同時作為冬季農田猛禽(紅隼、東方澤鵟、遊隼)的潛在獵物(被捕食者層次),在台灣低地農業景觀的冬季能量流動中承擔一個規模有限但功能確實的生態中介角色。其存在對農業生態系統整體功能的影響雖然難以精確量化,但其作為「健康農業生態指示物種」的潛力是存在的——大花鷚出現的農田,通常是昆蟲多樣性較高、農藥使用相對較低的農業生態友好型環境,其出現與否可以作為農田生態健康性的一個間接生物指標。
8.2 台灣越冬生態:棲地分布、族群密度與同域競爭種
大花鷚在台灣越冬期間的具體棲地分布,主要集中於以下幾個類型的開闊低地環境:宜蘭平原的廣大農田和濕地草地緣(宜蘭是台灣大花鷚記錄最穩定的地區之一);桃園大園一帶的農業田野(靠近桃園機場的低地農業區,是台北以北大花鷚越冬記錄的重要地點);台南和嘉義的西南部農業平原(尤其是收割後的稻田和魚塭草地緣);以及台北盆地周邊的河濱公園草皮地帶(如關渡自然公園周邊的草地)。
台灣大花鷚的越冬族群密度難以估計,由於其「不普遍」的定性,每年的出現總數量可能在數十至數百隻個體之間,但缺乏系統性的全島計數資料。從eBird台灣的年度記錄分佈來看,每年有記錄的大花鷚個體,以10月至12月之間的秋季記錄最為集中,其次是1月至2月的越冬期記錄,春季(3至4月)的過境記錄則相對較少。
與大花鷚同域越冬的鷚屬物種主要是樹鷚(Anthus hodgsoni,台灣最常見的越冬鷚類,偏好灌叢和林緣草地)和赤喉鷚(Anthus cervinus,常見冬候鳥,偏好短草地和農田),三者在棲地選擇上有一定的重疊但也有微棲地偏好的差異,形成一定程度的生態位分化(niche partitioning):大花鷚偏好最開闊、植被最矮短的地面環境;赤喉鷚能夠利用稍微茂密一些的短草地;樹鷚則更多出現在有灌木或稀疏樹木的草地邊緣。
8.3 台灣賞鳥文化中的大花鷚:鷚類辨識挑戰與鳥類學教育意義
在台灣的賞鳥文化中,大花鷚因其「不普遍冬候鳥」的身份和辨識難度,在一般入門賞鳥者的知識體系中屬於相對邊緣的物種,遠不如白鶺鴒、大卷尾或台灣藍鵲那樣為大眾所熟知。然而,在進階賞鳥者和鳥類學研究者的群體中,大花鷚及其相關的鷚類辨識課題具有相當高的討論熱度——鷚類辨識(pipit identification)在全球賞鳥圈中是公認的高難度技術領域,而在台灣能夠穩定記錄和辨識大花鷚(尤其是與布萊氏鷚的精確辨識),被視為鳥類辨識技術進階的重要里程碑之一。
大花鷚的鳥類學教育意義,體現在幾個面向:首先,其分類學歷史(從超種複合體到獨立物種的分裂過程)是理解現代鳥類系統學如何運作、以及「物種概念」在分類學實踐中如何應用的生動教材;其次,其後爪形態特徵和翼覆羽斑紋在辨識中的關鍵作用,訓練觀察者在田野中注意細節、發展精確形態觀察能力;第三,其連結西伯利亞繁殖地和台灣越冬地的長距離遷徙,為理解東亞候鳥遷徙網絡和台灣在這一網絡中的地位提供了具體的物種案例。
8.4 IUCN評估、棲地威脅與西伯利亞繁殖地的氣候變遷風險
大花鷚被IUCN紅皮書評定為「無危(Least Concern,LC)」,依據是其廣大的分布範圍(估計超過1000萬平方公里)、相對龐大的整體族群數量,以及目前尚未偵測到族群快速下降的跡象。然而,「無危」的當前評估並不意味著大花鷚在未來不面臨保育壓力,以下幾個潛在的長期威脅值得關注。
西伯利亞和蒙古草原的繁殖地棲地質量,可能受到氣候變遷驅動的植被帶移動(草原北移、灌叢化 shrubification)的長期影響,當低矮開放草地面積縮減、被灌木或樹木所取代時,大花鷚的繁殖棲地質量將隨之下降。蒙古草原的過度放牧問題,雖然短期內維持了低矮植被的開放景觀,但長期的土壤退化和植被組成改變,也可能對繁殖地的昆蟲多樣性和食物可獲性造成負面影響。在越冬地,農業集約化(intensive agriculture)導致的農藥使用增加、農田昆蟲多樣性下降,是影響大花鷚越冬地食物質量的潛在威脅,這一問題在印度和東南亞的農業景觀中已有不同程度的體現。
氣候變遷對大花鷚的影響,具有雙面性:一方面,如Dufour等人2021年研究所示,氣候變遷可能已在推動部分大花鷚族群建立全新的西向遷徙路線,顯示出這個物種在行為層面具有一定的氣候適應靈活性;另一方面,繁殖地草原的植被帶移動和越冬地農業生態系統的質量下降,可能在更長的時間尺度上對大花鷚的整體族群施加下行壓力。這種「短期行為適應性」與「長期棲地威脅」之間的張力,構成了大花鷚未來保育評估的核心不確定性。
8.5 未來研究課題:衛星追蹤、族群遺傳學與遷徙適應演化
大花鷚在科學研究層面,有幾個極具潛力的未來研究方向。首先,在台灣越冬個體身上進行的亞種鑑定和族群遺傳學研究,能夠精確確認台灣越冬族群的繁殖地來源,為建立台灣在東亞候鳥遷徙網絡中的具體角色提供數據支撐;通過線粒體DNA和核SNP(單核苷酸多態性)的分析,可以量化各亞種族群之間的遺傳分化程度,以及 sinensis 亞種與稻田鷚之間的基因邊界,對解決長達數十年的分類學爭議具有直接貢獻。
其次,在台灣越冬的大花鷚個體上安裝超輕型追蹤設備(如光定位器 geolocators,或更小型的GPS追蹤器),追蹤其北返後的繁殖地和遷徙路徑,能夠為建立「台灣—繁殖地」的完整遷徙連通性(migratory connectivity)數據提供直接證據,這不僅對大花鷚的生物學研究有價值,也對整個東亞鳥類遷徙網絡的保育規劃具有廣泛的參考意義。第三,深入研究大花鷚西向新遷徙路線的遺傳基礎(是遺傳決定的方向偏好突變,還是社會學習傳播的文化遷徙傳統?),對理解遷徙行為的演化機制具有根本性的科學意義,是目前鳥類遷徙行為進化生物學研究的最前沿課題之一。
總結
大花鷚的故事,是一個關於「距離」的故事。
牠在台灣冬季田野的出現,是一個跨越四千至六千公里長途跋涉的結果——從西伯利亞南部或蒙古草原的繁殖地,越過中國大陸,穿越台灣海峽,抵達台灣西部農田的土地。這段旅程在牠身上沒有留下任何外觀的痕跡,唯一可以看見的,是那個相比其他小型鳥類略顯誇張的筆直站姿,以及起飛時那一聲爆炸性的「shreep」——而正是這兩個特徵,讓有準備的觀察者能夠在田野的角落中辨認出這位遠道而來的草原旅人。
從命名學的起點回望,大花鷚的學名 Anthus richardi 承載了一段幾乎被歷史遺忘的人物故事:法國洛林省呂內維爾郵政局長查爾斯·理查(Charles Richard,1745—1835)在1818年前後收集到幾件大花鷚標本,轉交給法國鳥類學家維埃約,後者以其姓名為這個物種命名,留下了 richardi 這個拉丁文屬格後綴。在其後近兩百年間,「理查先生」的真實身份幾乎完全消失在文獻記載之中,直至2015年才被英國學者古拉德重新確認——這個小小的命名謎案,是科學知識建構過程中人物與名稱如何分離又如何重逢的生動縮影。
從演化的時間維度看,大花鷚屬於鷚屬在古北界草原的輻射擴散系譜,在更新世的氣候波動中,其祖先族群在西伯利亞至蒙古草原一帶逐步分化,形成了今日所見包含五至七個亞種的地理多樣性格局。台灣在大花鷚的分布體系中,處於越冬範圍的東緣,接收來自俄羅斯遠東和中國東北繁殖地的 ussuriensis 亞種為主的個體,是東亞候鳥南向遷徙主幹道的一個東側分支站點。
在外觀形態上,大花鷚的每一個形態細節都服務於其地面草地型生活方式:褐底黑紋的保護色系統提供視覺融合;超長而筆直的後爪提供地面行走的穩定支撐;強而有力的尖長翼型支撐長距離遷徙;爆炸性的「shreep」叫聲在起飛的瞬間穿透開闊空間傳遞存在信號。翼中覆羽末端的菱形尖銳深色斑,則是在近距離觀察中與布萊氏鷚最可靠的形態辨識依據。
在生理層面,大花鷚的遷徙前脂肪積累機制、遷徙期的器官可塑性(飛行肌增大、消化器官縮小),以及多模態整合的遷徙導航系統(地磁感知、太陽羅盤、星象定位和地標記憶的相互校正),構成了支撐數千公里長途遷徙的精密生理工程。這一生理架構在候鳥群體中高度保守,卻在大花鷚身上因遷徙距離的特別長途性而得到最充分的展示。
在行為生態層面,大花鷚筆直的直立站姿、緩步地面搜尋的覓食節律,以及面對天敵時以早期偵測加快速起飛為核心的防禦策略,共同構成了一個高度適應開闊無遮蔽草地環境的行為型,是與台灣藍鵲的林冠型行為系統和大卷尾的制高點守株型策略截然不同的第三種台灣鳥類行為生態面向。
在遷徙生態學的前沿,大花鷚的西向新遷徙路線,是21世紀最重要的鳥類遷徙行為演化案例之一。Dufour等人2021年在《當代生物學》發表的研究,首次在當代時間尺度上直接記錄到一個新遷徙路線正在鳥類族群中建立的過程,而氣候變遷被確認為最可能的驅動因素——這一發現不只是大花鷚的生物學突破,更是理解「動物行為如何在人類世(Anthropocene)快速演化」這一宏大課題的一個具體而深刻的窗口。
在保育生態學的框架中,大花鷚目前的保育壓力雖尚未達到緊迫的瀕危水準,但西伯利亞繁殖地的草原生態變遷和越冬地農業集約化的長期威脅,以及氣候變遷對其遷徙時序和路線的潛在干擾,是需要持續監測和研究的長期課題。台灣在這一保育框架中的角色,是作為東亞遷徙網絡的重要越冬站點,確保台灣低地農業景觀中的昆蟲多樣性和開闊棲地的連通性,為包括大花鷚在內的多種候鳥提供優質的越冬環境支持。
那個站在台灣冬季農田邊緣的褐色身影,以其不起眼的外表,在短暫停留的數月間,靜靜地連結著遠在西伯利亞草原的繁殖地、此刻的台灣田野,以及遙遠的演化歷史。當牠在一聲「shreep」中振翅離去,那道飛行的弧線,是草原給台灣送來的一封無聲的自然信件。
專業標籤(Tags)
#大花鷚 #理氏鷚 #Anthus_richardi #Richards_Pipit #鶺鴒科 #Motacillidae #鷚屬 #Anthus #雀形目 #Passeriformes #冬候鳥 #Winter_Visitor #不普遍冬候鳥 #台灣鳥類 #東古北界繁殖鳥 #長距離遷徙 #long_distance_migrant #西伯利亞繁殖地 #蒙古草原 #維埃約 #Vieillot #1818年命名 #查爾斯理查 #Charles_Richard #呂內維爾 #命名歷史 #亞種多樣性 #richardi亞種 #dauricus亞種 #centralasiae亞種 #ussuriensis亞種 #sinensis亞種 #超種複合體 #Anthus_novaeseelandiae #物種分裂 #布萊氏鷚 #Anthus_godlewskii #Blythspipit #稻田鷚 #Anthus_rufulus #後爪 #hindclaw #超長後爪 #筆直後爪 #直立站姿 #upright_posture #菱形深色斑 #median_wing_coverts #pointed_tip #shreep叫聲 #explosive_call #保護色羽衣 #褐色縱斑 #遷徙導航 #地磁感知 #magnetoreception #太陽羅盤 #星象定位 #脂肪積累 #pre_migratory_fattening #器官可塑性 #organ_plasticity #西向新遷徙路線 #westward_migration_route #當代生物學 #Current_Biology #Dufour_2021 #氣候變遷驅動 #climate_change_migration #先鋒個體 #pioneer_migrants #迷鳥到先鋒 #vagrancy_to_migration #光定位器 #geolocators #衛星追蹤 #東亞遷徙網絡 #East_Asian_flyway #宜蘭農田 #台灣越冬地 #農業昆蟲消耗 #生態服務 #IUCN_LC #草原生態系 #繁殖地保育 #氣候變遷風險 #鷚類辨識 #pipit_identification #台灣賞鳥 #進階辨識 #公民科學 #eBird #台灣生物多樣性 #候鳥保育 #東亞候鳥 #遷徙生態學 #migration_ecology #行為演化 #behavioral_evolution #遷徙適應 #migration_adaptation





















