第一章:物種概述
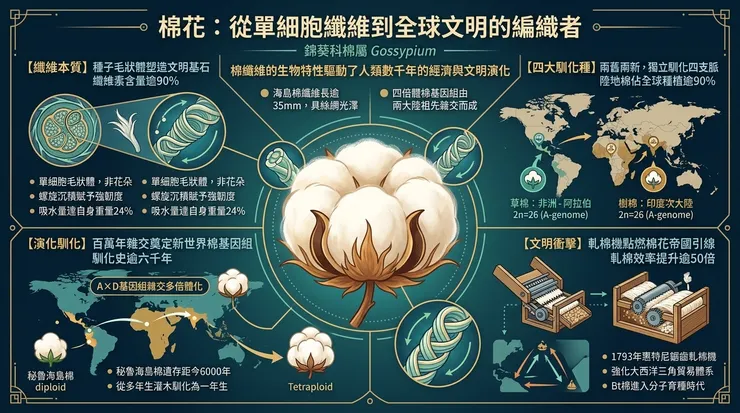
在人類文明的物質基礎中,少有如棉花這般,以如此柔韌的姿態,纏繞並塑造了全球的經濟版圖、社會結構與生態足跡。從植物學的嚴格定義而言,我們所珍視的「棉花」,並非花朵本身,而是錦葵科(Malvaceae)棉屬(*Gossypium*)植物種子上著生的單細胞毛狀體。這些纖維的主要成分是超過90%的純纖維素,以獨特的螺旋狀結構纏繞生長,長度可達數公分,其細胞壁在成熟後期次生加厚並脫水,形成中空的管狀結構,具備了強度、韌性與吸濕性的理想平衡。這一生物學上的偶然產物,因其物理化學特性,意外地與人類的技術需求耦合,開啟了一段跨越數千年的共演歷史。
棉屬包含約50個物種,其中僅有四個經歷了獨立且完整的馴化過程,成為今日全球紡織業的基石。這四個馴化種清晰地分為兩組:舊世界二倍體棉與新世界四倍體棉。舊世界棉包括起源於非洲-阿拉伯地區的草棉(*Gossypium herbaceum*)以及源於印度次大陸的樹棉(*Gossypium arboreum*)。兩者均擁有13對染色體(2n=26)。新世界棉則為異源四倍體,包含26對染色體(2n=52),其基因組被標記為(AD)n,是約一百萬至兩百萬年前,一個類似於舊世界A基因組的二倍體祖先與一個類似於新世界D基因組的二倍體祖先發生種間雜交,並伴隨基因組加倍事件而形成的。這兩個物種是陸地棉(*Gossypium hirsutum*),原產於中美洲及加勒比地區,以及海島棉(*Gossypium barbadense*),原產於南美洲安第斯地區及沿海。第一節:植物學本質與其「白色黃金」的由來
棉花的經濟價值核心,在於其種子纖維。從發育生物學角度看,棉纖維是從種子表皮細胞分化而來的單細胞結構。在開花授粉後約24小時內,種子表皮上特定細胞開始極化膨大,啟動纖維的初生壁合成。隨後進入為期約三週的次生壁合成期,纖維素以每日數微米的速度,圍繞中空腔(腔胞)呈Z形螺旋角(約20-30度)沉積,形成每日一層的生長輪。這種沉積方式賦予纖維獨特的機械性能:螺旋角較小時,纖維強度更高;角度較大時,則延展性更佳。纖維最終長度、強度與細度,由品種的遺傳潛力與生長期間的環境因子(水分、溫度、礦物質營養)共同決定。
纖維成熟的終點是「裂鈴」。蘋果狀的蒴果(棉鈴)果皮脫水後沿背縫線開裂,暴露出內部附著纖維的種子。此時,纖維細胞原生質體已程序性死亡並分解,殘留的細胞壁在日光與空氣作用下進一步脫水、扁平並扭曲,形成天然的可紡曲度。此曲度增加了纖維間的摩擦與抱合力,使其無需過度加撚即可紡成強韌的紗線。與動物纖維(如羊毛、絲)由蛋白質構成不同,棉纖維素的高度結晶性使其耐鹼性強,但易受強酸水解;其分子鏈上豐富的羥基使其親水性極佳,能吸收最高達自身重量24%的水分而不產生潮濕感,同時透過毛細作用促進水分蒸發。
海島棉(如著名的埃及長絨棉與秘魯皮馬棉)的纖維長度通常超過35毫米,且纖維細胞壁更薄、更光滑,賦予其絲綢般的光澤與超凡的柔軟度。陸地棉纖維較短(約25-30毫米),但單位面積產量高,適應性廣,佔據全球逾90%的商業種植面積。這種植物學特性上的分野,直接預示了它們在人類歷史中將走向不同的社會經濟軌跡:海島棉成為奢侈品與高端紡織品的原料,而陸地棉則註定推動大規模工業化生產。
演化地理與馴化路徑
棉屬的演化史是一部大陸漂移、氣候變遷與生物適應的編年史。分子系統學證據表明,棉屬的共同祖先可能起源於白堊紀的岡瓦納超大陸。隨著大陸分裂,其後裔在隔離的環境中分化。關鍵的演化事件發生在距今約一百萬至兩百萬年前,一個A基因組祖先(類似今日的非洲野生棉 *Gossypium herbaceum* 的變種 *africanum*)與一個D基因組祖先(類似今日的墨西哥野生棉 *Gossypium raimondii*)的雜交與多倍體化事件。這一事件可能發生在當時尚存陸橋連接的中美洲地區,創造了所有新世界四倍體棉的祖先。
馴化則是另一條獨立但並行的敘事線。舊世界棉的馴化發生在至少五千年前。草棉可能首先在非洲之角或阿拉伯南部被利用,隨後向東傳播至印度河流域。樹棉則在印度河流域文明(哈拉帕文化)中被獨立馴化,考古遺址中發現的棉織品殘片與棉籽印痕證實了這一點。新世界棉的馴化更早,考古學在秘魯北部海岸的華卡普列塔遺址發現了距今約六千年的海島棉纖維與紡錘,在墨西哥特瓦坎山谷發現了距今約五千五百年的陸地棉遺存。
馴化的標誌性性狀選擇包括:從多年生灌木向一年生習性轉變,以適應季節性農業;棉鈴從易脫落變為不易脫落,方便集中採收;種子從緊密附著纖維變為易於軋棉分離;最重要的是,纖維產量與品質的顯著提升。這些性狀的改變並非一蹴而就,而是在數千年的時間裡,透過人類無意識的選擇與後期有意識的育種,對棉株基因組施加的定向壓力。馴化過程深刻地改變了植物的生態位,使其從分散的野生群落,轉變為依賴人類播種、灌溉與保護的單一栽培體系。
從植物纖維到文明媒介
棉纖維的物理特性,使其成為前工業時代幾乎無可替代的紡織原料。與亞麻相比,它更柔軟、更易染色;與羊毛相比,它更輕盈、更適合溫暖氣候;與絲綢相比,它的生產不依賴難以規模化的昆蟲養殖。然而,將附著短絨的棉籽轉變為潔淨的纖維,需要耗費大量勞力。傳統的手工軋棉效率低下,這在長時段內限制了棉花生產的規模與地理擴張。
這一自然屬性造成的瓶頸,直到十八世紀末才被技術革新所突破。1793年伊萊·惠特尼的鋸齒軋棉機,透過簡單的機械原理——讓鋸齒滾筒穿過格柵,將纖維從棉籽上強力扯下——將軋棉效率提升了五十倍以上。這項發明直接強化了美國南部對奴隸勞動力的依賴,並將該地區與英國蘭開夏郡的工業紡織城鎮緊密連結,形成一個橫跨大西洋的「棉花帝國」三角貿易體系:美洲的土地與強制勞動、非洲的人口、歐洲的資本與工業製成品。
從更宏觀的文明史視角看,棉花作物的擴散與適應,反映了人類對不同生態區的改造能力。灌溉技術的發展使棉花得以在印度乾旱的德干高原、埃及的尼羅河河谷、美國的西部乾旱地區大規模種植。育種技術則不斷調整著物種的性狀,以抵抗病害(如棉鈴象鼻蟲、黃萎病)或適應機械化採收(培育果枝節位高、棉鈴集中吐絮的品種)。今日,經過基因改造的Bt棉(能產生蘇雲金桿菌毒素以對抗鱗翅目害蟲)已佔據全球主要棉區,這標誌著人類對棉花基因組的介入進入了分子層面。
因此,棉花不僅是一種作物,它是一個生物物理學上的介面,一頭連接著陽光、土壤、水與基因,另一頭連接著市場、工廠、資本與權力。其纖維的微觀結構,最終編織成了全球化的第一張巨網。理解棉花,便是理解自然物的屬性如何被納入人類的經濟與社會計算,並在漫長的時間中,反過來重塑了自然景觀與人類自身的命運。
第二章:演化與多樣性
棉花,作為人類衣著文明的基石,其故事始於數百萬年前的演化分歧與地理隔離。它並非單一物種的產物,而是一個屬(*Gossypium*)內多樣化策略的集合體。理解其生物學上的差異,是解讀其如何被不同文明擷取、並進而重塑文明軌跡的關鍵。本章將從棉屬的植物學溯源出發,剖析其主要栽培種的特徵與環境耐受性,最終勾勒其從野生祖先行散至全球農田的歷史路徑。
棉屬的系譜與起源中心
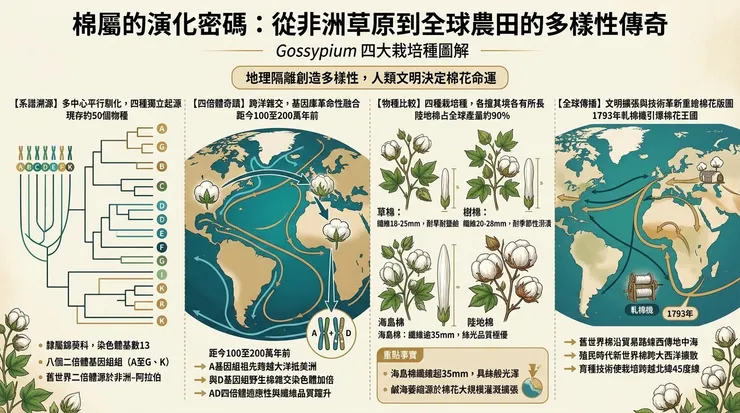
棉屬(*Gossypium*)隸屬於錦葵科(Malvaceae),該科植物以其韌皮纖維與特定的花部結構著稱。現存約50個棉屬物種,廣泛分布於全球熱帶與亞熱帶乾旱及半乾旱區域。透過細胞遺傳學分析,可將其劃分為八個二倍體基因組組(A至G,以及K),其染色體基數為13。其中,主宰全球棉花生產的,是四個獨立馴化的栽培種,它們分屬不同的基因組,揭示了多中心、平行馴化的歷史。
兩個舊世界栽培種——草棉(*Gossypium herbaceum* L., A₁基因組)與樹棉(*G. arboreum* L., A₂基因組)——均為二倍體(2n=26)。其野生祖先的遺傳證據指向非洲-阿拉伯板塊。具體而言,草棉的現存野生變種 *G. herbaceum var. africanum* 分布於非洲南部,顯示其馴化可能發生於非洲,隨後向北傳播至阿拉伯半島。樹棉的野生祖先形態已不復存在,但分子鐘推算與其密切相關的非洲野生二倍體物種的分化時間,暗示其馴化可能發生在印度次大陸,源自早期引入的A基因組祖先。
兩個新世界栽培種——陸地棉(*G. hirsutum* L., AD₁基因組)與海島棉(*G. barbadense* L., AD₂基因組)——則是異源四倍體(2n=52)。它們的形成是植物演化史上一次罕見的雜交事件:約在100萬至200萬年前,一個A基因組的二倍體祖先(與舊世界棉祖先相近)的種子,可能藉由洋流或鳥類,跨越了大西洋或太平洋的地理隔閡,與美洲的D基因組二倍體野生棉(如 *G. raimondii*)相遇並雜交,隨後染色體加倍,形成了全新的AD基因組物種。陸地棉的馴化中心在中美洲墨西哥一帶,而海島棉則在南美洲的安第斯山區及沿海地帶被獨立馴化。
這四倍體的形成,不僅是染色體數目的簡單疊加,更是一次基因庫的融合與創新,為棉花帶來了更強的適應潛力與更優異的纖維品質,預示了其未來征服全球紡織業的命運。
物種特徵與環境耐受譜系
四個栽培種在形態、農藝性狀及環境偏好上呈現顯著差異,這些差異直接源於其各自的演化背景與馴化選擇壓力。
草棉(*G. herbaceum*) 植株矮小,呈灌木狀,高度通常在一米以下。其棉鈴小,呈圓形,鈴殼厚,纖維短而粗,長度通常在18至25毫米之間。葉片具3至5個淺裂。它表現出極強的乾旱與高溫耐受性,是典型的短日照植物。其根系能有效利用貧瘠土壤中的水分,對土壤鹽鹼度也有一定忍耐力,這與其起源於非洲乾旱稀樹草原的環境密切相關。
樹棉(*G. arboreum*) 植株為多年生灌木,可達兩米以上,木質化程度高。棉鈴呈尖長的圓錐形,鈴殼較薄,纖維長度中等,約在20至28毫米,纖維強度較高。葉片裂刻較深。它適應亞熱帶季風氣候,能忍受季節性澇漬,但對持續低溫敏感。在印度,其栽培常與雨季同步,利用其較長的生長期。
陸地棉(*G. hirsutum*) 現代農業中占絕對主導(約90%產量)。其被馴化後選擇出一年生、緊湊、高產的類型。植株形態變異大,從矮叢到較高灌木皆有。棉鈴大,常為卵圓形,鈴殼薄,易於機械採收。纖維長度範圍廣(25-33毫米),細度與強度平衡良好。其最大的優勢在於廣泛的環境適應性:光周期反應較不敏感,使其緯度適應範圍大幅北擴;對多種土壤類型(從砂壤到黏土)有較好適應力;通過育種,其對主要病害(如黃萎病、枯萎病)及蟲害(棉鈴蟲)的抗性不斷增強。
海島棉(*G. barbadense*) 以其極品纖維著稱。植株生長勢強,分枝多。棉鈴較長,尖端顯著,通常三室或四室。其纖維特徵是極長(常超過35毫米)、極細、強度高且具有絲般光澤,這與其纖維細胞初生壁更薄、纖維素微纖絲排列更平行有關。然而,其代價是對環境要求嚴苛:需要長且溫暖無霜的生長期,對水分脅迫敏感,產量通常低於陸地棉。它主要局限於如埃及尼羅河谷、美國西南部、中國新疆等特殊灌溉農業區與海洋性氣候島嶼。
傳播路徑:生物潛能與人類選擇的交織
棉花的全球分布圖景,是其內在生物學特性與人類經濟活動、技術傳播及帝國擴張相互作用的結果。
舊世界棉的傳播路徑與早期農耕文明交疊。草棉從非洲之角向東,經阿拉伯半島傳入波斯、印度。樹棉在印度河流域文明(哈拉帕文化)中已有明確考古證據(約公元前2500年),其後向東傳至東南亞,向西至美索不達米亞與埃及。希臘與羅馬時代,棉花作為一種珍奇商品從東方引入地中海世界,但並未大規模栽培。
新世界棉的發現與傳播,則與地理大時代及殖民經濟直接掛鉤。哥倫布首次在美洲見到棉花。西班牙人將海島棉從秘魯帶回歐洲,並在殖民地試種。更具轉折意義的是陸地棉的全球化。18世紀末,北美南部殖民地大規模種植陸地棉,但其發展受制於手工分離棉籽的極高效率瓶頸。1793年惠特尼軋棉機的發明,使高效率處理短纖維陸地棉成為可能,直接引爆了美國的「棉花王國」時代,並深刻嵌入了跨大西洋的奴隸貿易與工業化紡織鏈條。
19世紀至20世紀,隨著育種技術(尤其是雜交與後來的基因工程)的進步,陸地棉的適應邊界被不斷推廣。例如,通過將海島棉的優質纖維基因導入陸地棉背景,培育出「皮馬棉」等長絨類型。同時,針對中國、中亞等溫帶地區,育成了早熟、耐低溫的品種,使棉花栽培跨越北緯45度線。灌溉技術的發展,則讓棉花得以深入如澳洲內陸、中亞沙漠等年降雨量不足250毫米的區域。
然而,這種人類驅動的擴張亦暴露並強化了棉花固有的環境弱點。其作為一種需水作物,在乾旱區的大規模單一種植導致了鹹海萎縮等生態災難;其對化學殺蟲劑的依賴,造成了土壤退化與生態系統失衡。棉花的多樣性,在人類追求單一經濟性狀的過程中,正面臨遺傳基礎狹窄化的風險。
綜上所述,棉花的演化與多樣性,是一部自然地理隔離創造遺傳原料、而人類文明進程進行篩選與強化的動態歷史。從非洲稀樹草原到美洲熱帶叢林,從二倍體到四倍體,從短粗纖維到極長絨,棉屬植物展示了生命應對乾旱、競爭與生殖的多元策略。而人類,則像一位專注的編輯,從這部浩瀚的基因文庫中,擷取出符合自身技術與經濟需求的章節,並將其書寫到全球地表,最終將一種區域性的野生植物,改造成為連結各大陸生態系統、經濟網絡與社會結構的全球性力量。其纖維所編織的,遠不止衣物,更是自然史與人類文明史緊密交纏的深層紋理。
第三章:外觀特徵
棉花,屬錦葵科(Malvaceae)棉屬(*Gossypium*),其當代形態是數千年自然演化壓力與人類定向選育共同作用的終端產物。它的外觀並非靜態的展示,而是一套針對特定生態區位——尤其是季節性乾旱與高溫環境——所發展出的生存方案。同時,每一處形態特徵的細節,都被人類農業實踐深刻閱讀、理解並重新詮釋,最終編碼入農耕文明的經濟紋理之中。本章將解剖棉株,從其物理結構的宏觀佈局到微觀解剖,揭示這些特徵背後的生態邏輯與歷史意涵。
整體形態與地下支撐:灌木的生存架構
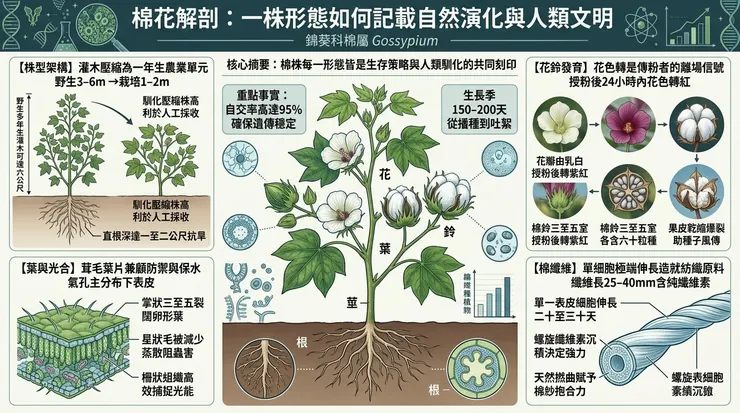
棉株在野生狀態下呈現典型的多年生灌木(shrub)或小喬木形態,這是在其起源地——乾熱的非洲、阿拉伯、澳洲及美洲等副熱帶地區——演化出的節能生長策略。其株高在未經馴化的品系中可達三至六公尺,形成多分枝的叢生結構,以增加光合作用表面積,同時在稀疏的林地或草原環境中有效競爭陽光。然而,在人類農業的介入下,特別是為便於手工採收而進行的選育,一年生栽培種的株高被大幅壓縮至一至二公尺範圍內,形態趨向緊湊,分枝節位降低,這是一項將植物生命史加速並置入年度耕作循環的關鍵馴化性狀。
其莖幹幼時呈綠色並具柔毛,隨生長進行初生與次生生長,逐漸木質化。莖的內部維管束排列呈現錦葵科典型的散生維管束(atactostele)特徵,提供機械支撐與輸導效率。分枝習性(determinate vs. indeterminate)是重要的農藝性狀:有限生長型(determinate)品種在主莖頂端直接形成花芽,停止向上生長,使成熟期一致,利於機械化收割;無限生長型(indeterminate)品種則持續由頂端分生組織產生營養結構,並從葉腋處形成花芽,開花結鈴期較長,在傳統農業中可應對不穩定的生長季。
支撐這地上架構的,是一套適應性根系。棉花發展出深達一至二公尺的直根系(taproot system),主根強健,能深入底土層汲取深層水分,這是對原生生境中季節性乾旱的直接適應。側根則廣泛水平延伸,形成密集的吸收網絡。在灌溉農業條件下,此根系模式能有效利用人工施加的水分與溶於水中的硝酸鹽、鉀離子等養分。然而,深耕與連作等農事操作,亦會改變土壤物理結構,影響根系分佈,進而反饋到地上部的生長勢與抗逆性。
能量捕獲與生殖結構:從葉片到棉鈴
棉花的葉片為互生,具長葉柄,葉片本身呈闊卵形或近圓形,通常具三至五個淺裂片(掌狀裂),裂片深度因品種而異。葉緣全緣,葉片兩面常被覆星狀毛或柔毛,此毛被層(pubescence)是一項重要的物理防禦與微氣候調節適應。毛被能增加葉片表面的邊界層厚度,減少水分蒸散;同時能對小型刺吸式昆蟲(如蚜蟲、葉蟬)的移動與取食造成物理阻礙,並影響其產卵選擇。葉片解剖顯示典型的雙面葉(bifacial leaf)結構:上表皮之下為緊密排列的柵狀組織(palisade mesophyll),富含葉綠體,負責高效的光合作用;下層為疏鬆的海綿組織(sporous mesophyll),利於氣體交換。氣孔(stomata)主要分佈於下表皮,通過調節其開閉來控制二氧化碳攝入與水分散失的平衡。
花器官是錦葵科的典型結構,具顯著的觀賞性,但此觀賞性實為繁殖策略的副產品。花單生於葉腋,具三枚大型的葉狀苞片(苞片通常深裂呈齒狀,稱為「副萼」),保護花蕾。花萼合生成杯狀,花瓣五枚,離生,初開時常呈乳白色或淡黃色,授粉後約24小時內因細胞液pH值變化及花青素(anthocyanin)的累積,轉為粉紅色或紫紅色,此色彩信號向傳粉者(主要為蜜蜂等昆蟲)表明該花已無訪問價值,從而提高訪花效率。雄蕊多數,花絲合生成單體雄蕊管(monadelphous stamens),包裹雌蕊的花柱。子房上位,由三至五心皮合生,形成未來棉鈴的雛形。
果實為蒴果,稱為「棉鈴」(boll)。棉鈴的發育始於授粉受精後,子房壁膨大形成堅韌、革質的外殼,內部空間被隔膜分隔成三至五室(locules),每室內著生六至十枚胚珠。棉鈴的成熟以「開裂」為標誌:沿著各心皮的背縫線(dorsal suture)與腹縫線(ventral suture),因纖維細胞的急劇膨脹產生內在張力,加上果皮組織的乾燥收縮,導致棉鈴爆裂,翻轉露出內部簇生的種子與纖維。此開裂機制對野生棉至關重要,確保種子能在乾季藉風力傳播。但在馴化過程中,人類傾向選擇開裂性較不完全或延遲的品種,以減少收穫前的損失。
棉纖維,即經濟產物,是單一表皮細胞經極端分化與伸長的結果。每一根纖維源自一個種子表皮細胞,在開花後約20至30天內,細胞以單一方向極性伸長,形成長度可達25至40毫米的細管(長絨棉品系更長)。此伸長階段結束後,細胞進入次生壁加厚期,以逆時針螺旋方式沉積幾乎純粹的纖維素(cellulose),螺旋角度影響纖維的強力與伸長率。纖維素沉積完成後,細胞原生質體死亡,留下中空的管狀結構,其橫截面呈腎形或腰圓形,具有天然的撚曲(convolution),此撚曲賦予棉紗抱合力與彈性。纖維表面覆有蠟質與果膠,需經由「精練」(scouring)等化學處理去除,方能獲得良好的吸濕性與染色性。
生命節奏與人類時序:從種子到田野
棉花的生活史特徵深刻體現了其對光、溫、水等環境因子的同步化適應。作為短日照植物(某些現代品種已培育為對光周期不敏感),野生與傳統品種在日照時間短於臨界值時,才從營養生長轉向生殖生長。這確保了開花與結鈴發生在溫度適宜、雨季末期的有利時段。從種子萌發到首次開花,約需60至90天,取決於積溫(accumulated growing degree days)。其生命週期在栽培條件下被嚴格壓縮為「一年生」,但在無霜凍的原生地,可作為多年生灌木存活數年,持續開花結果。
繁殖以自花授粉(self-pollination)為主,因雄蕊管包裹花柱,常於花冠開放前即已完成授粉,自交率可達95%以上。然而,其花器結構仍保留異花授粉(cross-pollination)的潛力,昆蟲訪花可導致約5%的異交率,這為種群維持了必要的遺傳多樣性,也為人類的雜交育種提供了生物學基礎。
種子的休眠(dormancy)與滯育(quiescence)是關鍵的生存策略。野生棉種子常具物理休眠(硬實種子,impermeable seed coat),需經過土壤微生物的分解、乾濕循環或通過動物消化道磨損種皮,方能打破休眠,確保在條件不利時不輕易萌發。栽培棉經過馴化,種皮通透性增強,休眠性減弱,以滿足人類對整齊、快速出苗的農藝要求。然而,種子仍能在乾燥、低溫條件下保持長達數年的活力(滯育狀態),這使得棉花種子成為早期貿易中可長期儲存、遠距離運輸的可靠商品。
人類農耕徹底重寫了棉花的生命劇本。生長期被嚴格定義在無霜期內,從播種到吐絮收穫約需150至200天。成年期——即具備生殖能力的階段——在人工栽培中始於首次開花,並通過持續的施肥、灌溉與病蟲害管理,盡可能延長其有效結鈴期,以最大化產量。其壽命在農業系統中僅為一個生長季,生命週期結束於機械或人工的採收行為。
農藝要點圍繞著對其形態與生理的精密調控:密植控制群體結構,影響通風透光與病蟲害發生率;灌溉模擬其原生境的降水模式,關鍵於花鈴期以減少蕾鈴脫落;施肥(特別是氮、磷、鉀的平衡)直接驅動營養生長與生殖生長的轉換;整枝(如去除徒長枝、抹贅芽)是人為導引光合同化物向棉鈴分配的干預措施;而化學調控(如施用縮節胺)則是直接干預其內源激素,抑制節間伸長,塑造理想的株型。這些操作,無一不是基於對棉花外觀特徵背後生理機制的解讀,是將野生植物的生存智慧,轉譯為大規模單一種植農業的生產指令。
因此,一株棉花的形態,從深扎的根系到爆裂的棉鈴,從茸毛的葉片到轉色的花瓣,都不僅是植物學的描述對象。它是一份由自然選擇起草,並經人類文明反覆批註與修改的複雜文本,記載著能量獲取、水分平衡、繁殖保障的古老法則,也銘刻著勞動組織、技術創新與全球貿易的近代歷史。
第四章:植物特性
棉花,作為人類文明史上最具影響力的經濟作物之一,其全球擴散的基礎,並非來自人類單方面的馴化意志,而是根植於其自身一系列複雜而精密的植物學特性。這些特性構成了它與環境對話的資本,也劃定了人類農業活動必須遵循的自然法則。本章將深入剖析棉屬植物的生存策略,揭示其如何在不同維度上,與陽光、水分、土壤及生物壓力進行協商,最終成為紡織工業無可替代的原料來源。
生存策略:環境耐受性與生長需求
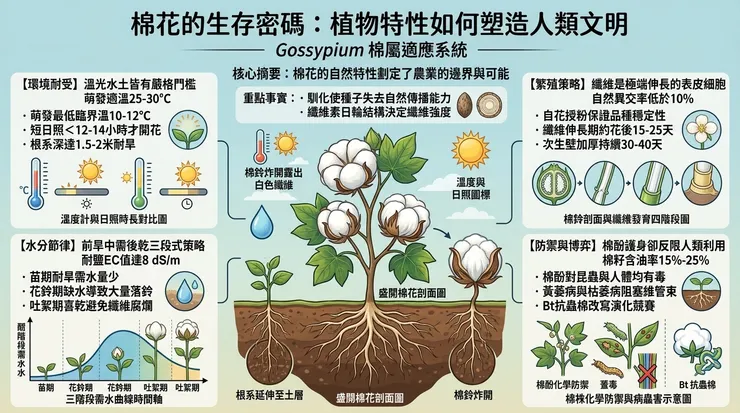
棉花屬(*Gossypium*)植物本質上是喜溫好光的多年生灌木或小喬木,在馴化過程中為適應一年生耕作而被塑造成一年生形態。其生長發育與環境因子存在著嚴格的量化關係。
溫度與光照:棉花是典型的喜溫與短日照植物。種子萌發的最低臨界溫度為10-12°C,最適溫度為25-30°C。從出苗到現蕾,需要有效的積溫(通常指≥10°C的溫度總和)。其光周期反應敏感,大多數栽培品種在每日光照時間少於12-14小時的條件下,才能從營養生長順利轉入生殖生長(現蕾、開花)。這一特性決定了其緯度分布界限,並在棉花從低緯度原產地向中緯度擴散時,成為人類必須通過選育早熟品種來克服的關鍵障礙。
水分與土壤:棉花被歸類為中等耐旱作物,但其需水規律呈現鮮明的階段性特徵。苗期需水量少,根系深扎可達1.5至2米,以利用深層土壤水。花鈴期(開花至棉鈴形成)是水分需求臨界期,此時水分脅迫將直接導致蕾鈴大量脫落。然而,成熟吐絮期則需乾燥環境,過多降雨會導致棉鈴腐爛、纖維品質下降。這種「前期耐旱、中期需水、後期喜乾」的水分需求模式,深刻影響了灌溉農業的發展與區域選擇。
土壤方面,棉花對鹽鹼和貧瘠的耐受性相對突出。其根系具有較強的離子選擇吸收能力,能在土壤電導率(EC值)達一定閾值(如8 dS/m)下維持生存。它偏好排水良好的砂質壤土,因其利於地溫回升與根系透氣,但通過人類的土壤改良措施,其適應範圍已被大幅拓寬。
繁殖策略:從花朵到纖維的形態學與生理學
棉花的繁殖過程是一場高度協調的形態建成與物質轉運的精密演出。
開花與授粉:棉花的花為兩性花,花瓣通常呈乳白色或黃色,開花當日即逐漸轉為粉紅色並凋謝,此顏色變化與花青素pH值改變有關。其花部結構(雄蕊管包裹雌蕊)有利於自花授粉,自然異交率通常低於10%,這保證了品種遺傳穩定性,是人類早期選育成功的基礎。然而,某些野生棉種或特殊品系仍保留著蟲媒傳粉的特徵,揭示了其演化歷史中的多樣化策略。
棉鈴與纖維發育:受精後,子房發育為蒴果,即棉鈴。棉纖維本質上是單一表皮細胞經極端伸長與次生壁加厚形成的植物毛。其發育可分為四個階段:起始、伸長、次生壁加厚與脫水成熟。在伸長期(開花後約15-25天),纖維細胞長度可達其最終長度的80%以上,此過程受內源激素(如生長素、赤黴素)精密調控。緊接的次生壁加厚期(約持續30-40天),纖維素以每日數層的速率向心沉積,形成日輪狀結構,此階段的持續時間與溫度直接決定了纖維的強度(纖維素含量與結晶度)。
種子與傳播:成熟棉鈴裂開(吐絮),露出被纖維包裹的種子。野生棉花的纖維短而稀疏,附著力強,易於藉助動物或風力傳播。馴化則強烈選擇了纖維長度、強度與著生密度,代價是種子失去自然傳播能力,完全依賴人類收穫與播種。棉籽含油率約15%-25%,蛋白質含量豐富,但同時含有對單胃動物有毒的棉酚,這一人類與棉花共演化的副產品,催生了後續的脫毒加工產業。
防禦、共生與人類的介入
在自然生態位中,棉花並非被動的資源提供者,它擁有一套化學與物理防禦體系,以應對植食性動物與病原菌的壓力。而人類的農業實踐,則是一場對這些自然特性進行持續管理、強化或規避的漫長對話。
化學防禦與毒性:棉株全身,特別是種子腺體,富含一系列次生代謝物,其中最著名的是棉酚,一種多酚類化合物。棉酚對昆蟲和軟體動物具有顯著毒性,是棉花天然的蟲害防禦機制。然而,它對非反芻動物(包括人類)同樣有毒,能與蛋白質結合、干擾代謝並可能導致組織損傷。棉籽的食用必須經過複雜的熱處理與溶劑萃取以降低棉酚含量。這一毒性特質,在歷史上既限制了棉籽的利用,也促進了現代油脂加工技術的發展。
病蟲害壓力:棉花是眾多病蟲害的寄主。關鍵病害包括由真菌引起的黃萎病(*Verticillium dahliae*)和枯萎病(*Fusarium oxysporum* f. sp. *vasinfectum*),它們通過土壤傳播,阻塞維管束,導致植株萎蔫死亡。蟲害方面,棉鈴象鼻蟲、棉蚜、棉鈴蟲等是主要威脅。這些生物壓力與棉花的大規模單一種植形成正反饋,迫使農業系統不斷回應:從輪作倒茬、藥劑噴灑,到培育抗病蟲的轉基因品種(如Bt抗蟲棉),人類的介入已深度改變了棉花與其天敵間的演化競賽。
抗逆性與農業邊界:棉花的抗風性一般,其較大的葉片和果枝結構在強風下易受機械損傷,導致落蕾落鈴。抗旱性則如前所述,其中等偏強,但極端乾旱仍會造成嚴重減產。抗鹽性是其得以在邊際土地(如中亞、美國西部部分地區)種植的關鍵,其耐受機制涉及滲透調節、離子區隔化與活性氧清除等複雜生理過程。人類通過灌溉洗鹽、選育耐鹽品種,不斷試探著棉花生長的生態邊界。
綜上所述,棉花的植物特性並非一組孤立的性狀列表,而是一個動態的、與環境緊密耦合的適應系統。它的每一項需求與耐受,都像一道無形的等高線,在地球表面劃出了可能的栽培區域。它的每一次開花、結鈴與纖維沉積,都是一系列嚴謹的生化反應在特定環境條件下的產物。人類文明對棉花的依賴,實質上是對這一套自然法則的深刻理解、巧妙利用與持續改造。從選擇適宜的日照與積溫地帶,到管理其獨特的水分需求曲線,再到與其病蟲害及自身毒性進行漫長博弈,農業史與紡織工業史的背後,始終貫穿著一部人類與棉花植物特性不斷協商的自然史。
第五章:棉花與人類的關係
棉花與人類的關係,遠超乎一種纖維作物與其使用者的簡單定義。這是一種由基因組、生態位、能量流動與全球貿易網絡共同編織的共生與衝突史。人類將野生棉屬植物納入其農業生態系統,徹底改變了物種的分布、遺傳多樣性與演化壓力;與此同時,棉花的生物學特性也反過來重塑了人類的經濟組織、技術發展與社會結構。本章將檢視棉花在自然食物網中的位置,並深入剖析人類對其進行系統性利用的複雜歷程,以及這種利用所引發的可持續性挑戰。
生態系中的消費者與被消費者
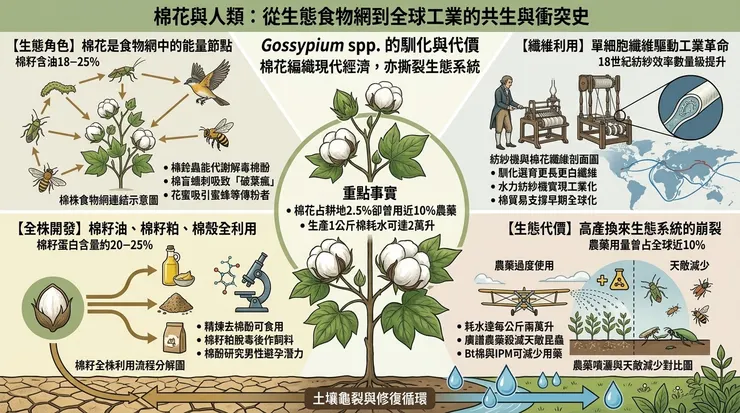
在人類大規模介入之前,棉花作為一種多年生灌木或小喬木,已是特定生態系中能量流轉的一環。其各組織部位為多種無脊椎動物及部分脊椎動物提供營養來源。
棉花的營養器官(葉片、嫩莖) 與生殖器官(花蕾、棉鈴、種子) 富含碳水化合物、蛋白質及油脂,吸引專化或泛化的植食者。最具代表性的專化性昆蟲是棉鈴蟲(Helicoverpa armigera) 及其近緣種。這類鱗翅目昆蟲的幼蟲具高度適應性的口器,能直接鑽入棉鈴內部取食發育中的纖維與種子。其消化系統能有效分解棉花組織中含有的棉酚(gossypol)——一種由棉株合成、用以抵禦病蟲害的萜烯類化合物。棉鈴蟲體內細胞色素P450單加氧酶系統的特定同功酶,能對棉酚進行代謝解毒,此為長期協同演化形成的抗性機制。此外,棉盲蝽(Apolygus lucorum) 等半翅目昆蟲則以刺吸式口器刺穿棉株幼嫩組織的韌皮部,吸取汁液,同時注入唾液導致細胞異常分裂,形成俗稱「破葉瘋」的結構性傷害。
棉花的花蜜與花粉為多種傳粉者,包括蜜蜂、蝴蝶及某些蠅類,提供能量與蛋白質來源。然而,現代大面積單一栽培的棉田,因其開花期集中且農藥使用頻繁,往往成為傳粉昆蟲的「生態陷阱」——提供短期食物,卻伴隨高死亡率風險。
在脊椎動物層面,棉籽因其高油脂含量(約18-25%)及蛋白質(約20-25%),成為鳥類(如野鴿、麻雀)及囓齒類動物(如田鼠)的覓食目標。棉花葉片偶爾也會被大型草食哺乳動物,如鹿或野羊取食,但這並非其主要食物來源。這些取食關係在自然狀態下構成動態平衡,但當棉花作為高密度單一作物存在時,特定植食者種群便可能爆發,從自然消費者轉變為農業意義上的「害蟲」。
馴化、利用與全球經濟體的塑造
人類對棉花的利用,是一場對其生物屬性進行全方位開發與改造的漫長歷程。此過程始於數千年前的獨立馴化事件,並隨著技術革新而不斷深化對該植物資源的榨取效率。
1. 纖維利用與紡織革命
棉花最主要的應用價值在於其種子表皮細胞特化而成的單細胞纖維。這些纖維初生壁以幾乎純粹的纖維素構成,在種子發育過程中次生壁以每日數微米的速度呈螺旋狀沉積加厚,形成中空結構,賦予其輕盈、透氣、強韌且易於染色的物理特性。人類的馴化選擇方向明確:增加纖維長度(絨長)、提高纖維強度、消除天然色素(培育出白色棉絮),並促使棉鈴開裂(吐絮)更為集中以便採收。
從手工摘棉、軋棉去籽,到紡紗、織布,棉花加工鏈驅動了連續的技術創新。18世紀英國的水力紡紗機與騾機,本質上是將棉花纖維的物理特性與機械動力結合,實現纖維拉伸、加撚與卷繞的工業化,導致紡紗效率呈數量級提升。隨後,棉紡織業成為工業革命的先導部門,其對原棉的龐大需求,直接強化了美洲的奴隸種植園經濟,並將印度等傳統產區重塑為原料供應地與商品傾銷市場。棉花貿易網絡成為早期全球化的骨架,其纖維不僅編織衣物,更編織了跨大陸的資本、勞動力與權力關係。
2. 種子副產品的綜合開發
棉籽的利用體現了對生物質的「全株利用」思維。棉籽含油量高,通過壓榨或溶劑萃取可得棉籽油。精煉過程需去除棉酚,使其成為安全的食用植物油,富含亞油酸。榨油後的殘渣即棉籽粕,蛋白質含量豐富,但同樣需經脫毒處理(如熱處理、化學萃取)以降低棉酚至安全標準以下,方能作為反芻動物及單胃動物的飼料蛋白來源。棉籽殼堅硬,主要用作培養基質或低熱值燃料。
3. 傳統與潛在的藥用價值
棉酚作為棉株的天然防禦化合物,在藥理學研究中被探討其潛在的生物活性。歷史上,棉根皮曾用於民間療法。現代研究聚焦於棉酚對特定細胞代謝路徑的干擾作用,例如其作為男性避孕藥的潛力(通過抑制精子發生),以及對某些癌細胞系的增殖抑制效果。然而,其全身性毒性(特別是對心臟與肝臟的損傷風險)及治療窗口狹窄,限制了其臨床應用。目前研究更傾向於對其化學結構進行修飾,以降低毒性、提高靶向性。
養殖難度主要源於其生物學弱點與集約化農業的內在矛盾。棉花對水分脅迫敏感,花期與鈴期需水規律,但又不耐漬澇。它對多種病蟲害缺乏抵抗力,尤其是黃萎病(Verticillium dahliae) 等土傳病害,以及棉鈴蟲、棉蚜等蟲害。這導致傳統棉花種植高度依賴灌溉、化肥與農用化學品(殺蟲劑、殺菌劑) 的投入。其生產構成一個典型的能量密集型系統,環境外部性高昂。
生態代價與可持續性路徑的探索
20世紀以來的集約化棉花生產模式,在提升單產的同時,也暴露了其生態與社會的不可持續性。其核心矛盾在於:為滿足單一物種(人類對纖維的需求)的最大化產出,而簡化乃至破壞了支持該物種生長的農業生態系統。
環境代價具體而微。長期單一連作導致土壤微生物群落失衡、有機質耗竭、次生鹽鹼化。為控制蟲害而大量使用的廣譜性合成殺蟲劑,在殺滅目標害蟲的同時,也消滅了其天敵(如瓢蟲、草蛉、寄生蜂),破壞了自然的生物控制機制,反而可能引發次級害蟲爆發。農藥與化肥的徑流與滲漏,對水體造成富營養化與污染。據聯合國糧農組織(FAO)與國際棉花諮詢委員會(ICAC)的數據,傳統棉花種植面積約占全球耕地的2.5%,但其農藥使用量曾長期占全球總量的近10%(隨著生物技術棉花的推廣,此比例已下降,但問題依然存在)。此外,棉花是極耗水的作物,生產一公斤原棉需耗水數千至兩萬升不等,取決於氣候與灌溉效率,這在乾旱與半乾旱產區加劇了水資源壓力。
面對這些挑戰,可持續利用策略正從多個層面展開:
1. 遺傳改良與綜合管理:推廣抗蟲轉基因棉花(如Bt棉) 能針對性控制鱗翅目害蟲,顯著減少殺蟲劑噴施次數。培育抗旱、耐鹽、抗病的傳統品種與新技術品種,以適應環境脅迫。結合害蟲綜合治理(IPM),利用性信息素干擾、釋放天敵、種植誘集作物等生態方法,將化學防治作為最後手段。
2. 有機與再生農業實踐:有機棉花種植嚴格禁止合成農藥與化肥,依賴作物輪作(與豆科、禾本科作物輪作)、種植綠肥、施用堆肥、以及使用生物源農藥來維持地力與控制病蟲害。其養殖難度體現在初期轉換期產量可能下降,病蟲害管理更依賴經驗與勞動力投入,但長期有助於恢復土壤健康與生物多樣性。再生農業則更進一步,強調通過免耕或少耕、持續土壤覆蓋、多樣化種植等措施,提升農田的碳匯能力與生態韌性。
3. 資源循環與認證體系:推動棉花副產物(棉籽、棉稈)的高值化利用,發展循環經濟。例如,棉稈可用於生產生物質能源或環保材料。同時,「良好棉花」(Better Cotton) 等全球認證體系,通過制定並推廣關於水資源管理、農藥合理使用、土壤健康與勞動權益的生產標準,引導大規模生產向更可持續的方向轉變。
棉花與人類的關係史,是一部從區域性生態適應到全球性工業剝削,再逐步轉向尋求生態平衡的漫長演變。這種植物的纖維編織了現代世界的經濟圖景,而其種植所引發的環境與社會問題,也迫使我們重新審視農業生產的底層邏輯。未來的路徑,不在於放棄利用,而在於如何依據其生物學本性,設計出一個能同時維繫生態系統服務、農民生計與纖維需求的、更具韌性的生產系統。這要求我們不僅將棉花視為一種商品,更應將其理解為一個需要與之協同共生的生命體。
第六章:棉花產業與文明的故事
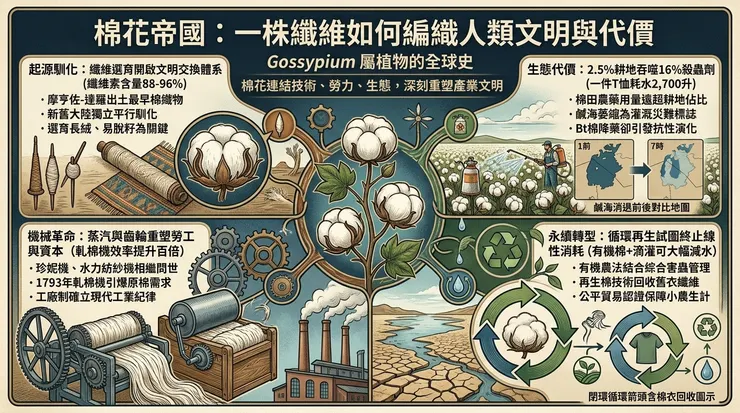
棉花,*Gossypium* 屬植物種子表皮衍生的單細胞纖維,其產業化歷程是人類將一種植物形態適應,轉化為全球性經濟與權力網絡的經典範例。這並非一個單純的技術進步敘事,而是植物基因、機械工程、能量來源、勞工組織與資本流動共同編織的深層結構史。
從野生棉鈴到全球商品鏈的開端
棉屬植物的演化路徑賦予了其被大規模利用的潛力。其蒴果(棉鈴)的裂開特性(裂蒴性)是一種種子傳播機制,但附著於種子表面的纖維——由表皮細胞突起、延長並加厚細胞壁(主要成分為約88-96%的纖維素)而成——則成為人類選擇的關鍵性狀。早期馴化聚焦於增加纖維長度(絨長)、強度與著生密度,並減輕種子與纖維的附著力,便於後續加工。
考古證據將人類系統化利用棉花的時間點置於數千年前。印度河谷文明的摩亨佐-達羅遺址(約公元前3250-2750年)出土了棉織物殘片;同時期,秘魯海岸的社會也獨立馴化了美洲棉種。這標誌著棉花在舊大陸與新大陸的文明中,同步從一種區域性資源,開始嵌入地方性生產與交換體系。其加工最初完全依賴人力:手工摘除棉籽(軋棉)、使用紡錘或原始紡車將纖維加撚成紗、再以腰機或踏板織機織造。這一階段的棉花,其流通半徑受限於原料重量與價值比,以及紡織所需的密集勞力投入,本質上仍是一種「奢侈品」或區域特產品。
然而,棉花的生物特性預示了其產業潛能:相較於羊毛,其纖維更易於均質化處理;相較於亞麻,其加工流程(從原棉到紗線)更為簡化。當它與遠距離貿易網絡和新型生產組織相遇時,變革的條件便告成熟。
機械能、勞工與生產關係的重塑
十八世紀後期至十九世紀,一系列技術創新系統性地突破了棉花產業的瓶頸,其核心是以非生物能量(水力、蒸汽)替代人力,並以機械裝置模擬並放大特定手工動作。這並非單一發明,而是一套相互鎖扣的技術體系。
首先是紡織環節的機械化。詹姆斯·哈格里夫斯的珍妮紡紗機(約1764年)增加了紗錠數量;理查·阿克賴特的水力紡紗機(1769年)利用水車動力生產出更堅韌的紗線,並確立了集中化工廠生產模式;塞繆爾·克隆普頓的騾機(1779年)結合兩者優點,能紡出既細且韌的優質棉紗。織布環節隨後跟上,埃德蒙·卡特賴特的動力織機(1785年)最終實現了織布的機械化。這些發明大幅提升了從纖維到布匹的轉化效率,但創造了一個尖銳矛盾:對原棉原料的饑渴式需求。
原棉處理的關鍵瓶頸在於軋棉——將纖維與種子分離。手工軋棉一人一日僅能處理約一磅。伊萊·惠特尼於1793年發明的軋棉機(儘管其專利爭議重重),通過一系列帶鉤齒的滾筒與固定柵條的相互作用,使日處理量躍升至數百磅。這項看似單一的機械革新,產生了深遠的結構性後果:它使大規模種植短絨陸地棉(*Gossypium hirsutum*)在美國南部變得極度有利可圖,從而強化了對奴隸勞動力的依賴。軋棉機解放了生產力,卻同時鎖死了種植園的社會生產關係。
至此,一個橫跨大陸的產業鏈閉環形成:美國南部(後來的全球多個產區)提供土地密集、勞力密集的原棉;英國蘭開夏郡等地提供資本密集、能源密集的紡織工廠;而全球市場,特別是殖民地和半殖民地,則成為其製成品的傾銷地。棉花成為「工業革命」的先行產業,其工廠制度確立了現代工業時間紀律與勞工管理範式。
生態足跡、基因改造與可持續性困境
二十世紀以降,棉花產業在農業與工業兩端持續演化,其影響深入生態與社會肌理。
在農業端,二戰後的「綠色革命」模式侵入棉田:高產品種需搭配密集灌溉、合成化肥與化學農藥。棉花種植面積約佔全球耕地的2.5%,卻消耗了約16%的殺蟲劑和約6-8%的化肥,其生態足跡顯著。例如,中亞的鹹海因大規模棉花灌溉而急遽萎縮,成為環境災難的標誌。為對抗鱗翅目害蟲(如棉鈴蟲),科學家將蘇雲金桿菌(*Bacillus thuringiensis*, Bt)中產生特定毒蛋白的基因轉入棉花,創造了Bt棉。這項技術在特定區域內顯著降低了殺蟲劑使用量,但亦引發了次生害蟲崛起、靶標害蟲抗性演化等生態學後果,並將農民置於對商業化種子與配套農藝的依賴之中。
在工業與消費端,全球化分工使紡織製造業從早期工業化國家向勞動力成本更低的區域轉移,重演了歷史上對勞力與環境成本的追逐。快時尚商業模式加劇了資源消耗與廢棄物問題,一件標準棉T恤的生產約消耗2,700升水(主要用於灌溉階段)。
面對這些困境,可持續策略在多層次展開:
1. 農業生態學實踐:推廣有機棉花種植,通過作物輪作、間作、釋放天敵等綜合害蟲管理策略,減少化學投入。採用滴灌等精準灌溉技術,降低水資源消耗。
2. 育種技術革新:除轉基因技術外,利用分子標記輔助選擇等技術,培育抗旱、耐鹽、纖維品質更優的傳統品種。
3. 循環經濟模型:發展棉織品回收技術,將舊衣物中的棉纖維經機械或化學法分解後,重新紡製成紗線(再生棉),減少對原棉的開採壓力。
4. 生產者與消費者責任:從公平貿易認證確保小農生計,到建立紡織品生命週期評估與標籤制度,試圖將環境與社會成本重新內部化於商品鏈之中。
棉花的故事,從一株熱帶灌木的纖維開始,最終編織成一部關於人類如何利用自然、重組社會、並不得不面對其行為所引發的複雜反饋的編年史。它揭示了產業文明的核心邏輯:通過技術與組織創新將地理上分散的資源、能量與勞力整合進單一系統,以追求效率與增長,而這一過程必然在自然生態與人類社會中同時刻下其深淺不一的痕跡。
