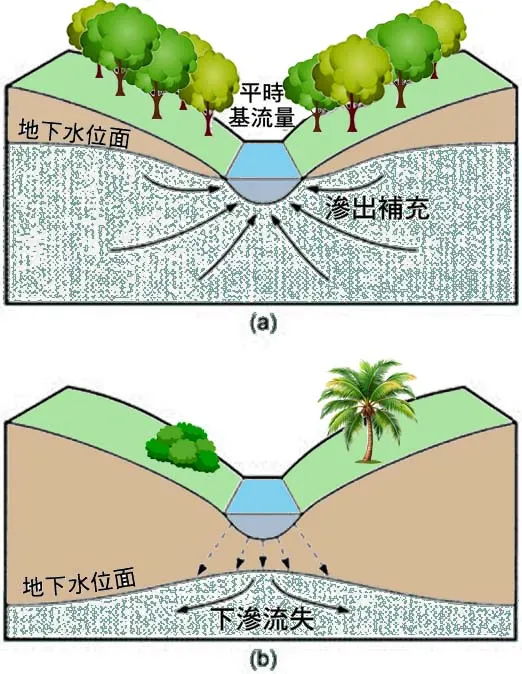
2.1 緩流區的沉積作用
緊隨在喧鬧的急瀨之後,河流常常會進入一段水面平靜、流動相對均勻的區域,這就是緩流區(Run),有時也稱為緩瀨。它在地形上通常比急瀨更深、坡度更平緩,是從急瀨的能量釋放區到深潭的能量儲存區之間的一個過渡地帶。如果說急瀨是河流的「衝刺賽道」,那麼緩流區就像是賽道旁的「緩衝區」和「能量轉移帶」。在這裡,水流的能量有所減弱,開始發生初步的沉積作用,塑造出與急瀨截然不同的物理環境和生態功能。
緩流區最核心的物理特徵是相對均勻且中等速度的水流。水流不再像急瀨那樣湍急翻滾,而是以一種較為層流(laminar flow)的方式向前推進。這種流態的能量恰到好處:它依然有足夠的力量將最細小的淤泥和黏土懸浮在水中並帶向下游,但已經無力推動較大的沙粒和細礫。因此,緩流區成為了中等粒徑顆粒的沉積區。其河床底質通常由較小的礫石和粗沙組成,顆粒大小明顯小於急瀨的卵石,但又比深潭的淤泥要粗。這種底質的孔隙度依然良好,能夠為底棲生物提供棲息空間,但其穩定性不如急瀨的巨石,在洪水期間可能會發生移動。
由於流速減慢,緩流區的再曝氣效率遠低於急瀨。然而,因為它緊接在急瀨之後,通常能接收到來自上游富含氧氣的水流,所以其溶氧量依然可以維持在一個相對較高的水平,足以支持大多數水生生物的生存。水深介於淺灘和深潭之間,陽光依然能夠照射到底部,支持附著藻類的生長,但其生產力可能略低於光照更充足的急瀨。
緩流區最重要的生態功能之一,是作為物質和生物的輸送通道與分揀區。從急瀨的生物群落中被意外沖刷下來的無脊椎動物,會在水流中形成一種生態學上稱為「漂流」(drift)的現象。這些漂流的昆蟲幼蟲,在進入流速減緩的緩流區後,一部分會重新找到合適的底質附著下來,實現了族群的向下游擴散;而另一大部分則成為了魚類唾手可得的食物。緩流區是漂流捕食性魚類(drift-feeding fish)最主要的覓食場所。魚類會選擇一個有利的位置,逆流懸停,像在自助餐的傳送帶前一樣,等待著從上游急瀨源源不斷「漂」來的食物。這種高效的覓食策略,使得緩流區常常聚集了大量的魚類。
此外,緩流區的沉積作用也為特定生物創造了棲地。中等粒徑的礫石和粗沙混合底質,是許多魚類理想的產卵場。例如,一些鯉科魚類會選擇在緩流區的礫石灘中產卵。同時,這種底質也適合一些需要部分鑽穴的無脊椎動物生存,例如某些蚌類和大型的蜻蛉稚蟲。相較於急瀨,緩流區的物理壓力較小,使得一些游泳能力較強、但固著能力較弱的生物也能在此棲息活動。
在整個「急流-緩流-深潭」序列中,緩流區扮演著一個承上啟下的關鍵角色。它將急瀨生產的氧氣和食物向下游輸送,並通過初步的沉積作用,開始篩選和重塑河床的樣貌。它不像急瀨那樣充滿能量,也不像深潭那樣沉靜穩定,但正是這種「中庸」的特性,使其成為一個物種來來往往、能量高效轉移的動態走廊。許多魚類可能會在深潭中休息和躲避天敵,在急瀨中尋找特定食物,但它們一天中大部分的覓食時間,可能都是在緩流區度過的。因此,一個功能健全的緩流區,對於連接不同棲地單元、維持魚類族群的健康至關重要。在河道整治工程中,常常會將河道渠化,形成均一的緩流,這雖然看似平穩,卻因為消滅了急瀨和深潭,極大地破壞了棲地的多樣性,最終導致生物群落的單一化和衰退。
2.1.3 深潭的形成機制
深潭(Pool),是河流中那些水深較深、水流異常平緩、甚至看似靜止的區域。它如同河流的「休憩室」與「避難所」,在喧囂的急流與平穩的緩流之間,提供了一個沉靜而穩定的世界。深潭的存在,極大地豐富了河流的垂直空間結構,為大型魚類、不同季節的生物以及躲避極端環境的物種提供了不可或缺的關鍵棲地。深潭並非隨機挖成,其形成機制與水流的能量轉換和河道的形態密切相關。
深潭的形成,本質上是水流侵蝕能量在特定地點集中的結果。一個常見的形成機制與河道的彎曲有關,即曲流(meander)的形成。在彎曲的河道中,由於慣性,水流會傾向於衝向凹岸(外側岸),並在凹岸下方形成一股螺旋狀的次級水流(helical flow)。這股水流會持續地掏蝕凹岸的根基,將泥沙挖走並帶到下游。與此同時,在凸岸(內側岸),流速則顯著減慢,導致從上游攜帶來的泥沙在此沉積,形成邊灘。經過長期的「凹岸侵蝕、凸岸堆積」作用,河道會變得越來越彎曲,而在被持續掏空的凹岸下方,便會形成一個狹長而深邃的深潭。
另一種常見的形成機制與河道中的障礙物有關。當水流遇到巨大的、難以動搖的障礙物時,如巨大的漂石、基岩出露,或是倒伏在河中的巨大樹木(large woody debris),水流會被迫繞過或越過障礙物。在障礙物的下游側,水流會形成強烈的渦流和向下的沖刷力,這種局部增強的侵蝕力量會不斷地挖掘河床,形成一個坑洞,即「沖刷潭」(scour pool)。倒伏木在形成深潭的過程中扮演著尤其重要的角色,它不僅能引發沖刷,其本身也為魚類提供了極佳的掩蔽結構,因此在河流生態修復中,常常會人為地放置大型木結構來營造深潭棲地。
此外,深潭也常常與急瀨成對出現,構成階梯-深潭(step-pool)系統,這在上游陡峭的溪流中尤為常見。在這種結構中,一排由大石頭構成的、類似微型瀑布的「階梯」(即急瀨)會攔截水流,水流在跌落階梯後,其勢能轉化為動能,在階梯的下游形成一個具有強大沖刷力的「跌水潭」(plunge pool)。這個潭會不斷地被加深,直到水流能量耗散,然後在下游不遠處,又會形成下一個由礫石堆積而成的階梯,如此循環往復,構成了階梯狀的河床縱剖面。
無論形成機制如何,深潭都具有一些共同的物理和生態特徵。其極緩慢的流速是其最顯著的特點。這使得即便是最細微的顆粒,如淤泥和有機碎屑,也能在此沉降下來。因此,深潭的底質通常是柔軟的淤泥或細沙,與急瀨的礫石底質形成鮮明對比。這種環境不適合固著生物,但卻是許多穴居無脊椎動物(如顫蚓、搖蚊幼蟲)的樂園。它們在富含有機質的底泥中覓食,扮演著分解者的重要角色。
由於水深較深,陽光難以穿透到底層,加上底泥中有機物分解會消耗大量氧氣,深潭底層的溶氧量通常較低,水溫也相對穩定。在夏季,當淺水區的水溫因暴曬而急劇升高時,深潭的底層依然能保持相對涼爽,成為許多魚類躲避高溫的「夏日避暑山莊」。反之,在冬季,當淺水結冰或水溫極低時,深潭較大的水體能更好地保溫,為魚類提供了越冬的場所。因此,深潭是河流中的「溫度避難所」(thermal refuge)。
深潭最重要的生態功能,是為大型魚類提供核心棲地。大型魚類需要足夠的活動空間和深度來躲避陸地上的天敵(如鳥類和哺乳動物)。深潭提供的深度和通常伴隨存在的掩蔽物(如倒木、深切的河岸)為它們提供了絕佳的安全性。它們會以深潭為基地,在一天中的特定時間(如晨昏)游到鄰近的緩流或急瀨去覓食,然後再回到深潭中休息和消化。可以說,一個河段能供養多大的魚,很大程度上取決於其深潭的體積和品質。
總結來說,深潭是河流棲地多樣性的關鍵一環。它通過多種水流力學機制形成,提供了與急流和緩流截然不同的深、緩、靜的環境。它不僅是大型魚類的家園,更是整個河流生物群落在面對季節性溫度壓力或枯水期棲地縮減時的生命庇-護所。一條失去了深潭的河流,就如同一個失去了臥室和庇護所的家園,其生命的承載能力和抵禦干擾的韌性都將大打折扣。
2.1.4 邊灘與河心洲的生態意義
在河流蜿蜒的旅程中,除了水下的急流與深潭,那些在洪水期被淹沒、枯水期則露出水面的沉積地貌——邊灘(Point Bars)和河心洲(Mid-channel Bars),同樣是構成河流生態系統多樣性的關鍵組件。它們是河流侵蝕與堆積作用達到動態平衡的產物,是河流生命力在陸地上的延伸。這些看似荒蕪的沙礫灘,實際上是充滿生機的演替前線、重要的生物棲地以及水陸交界帶的生態緩衝區,具有不可忽視的生態意義。
邊灘,主要形成於彎曲河道的凸岸(內側岸)。正如前述,水流在凹岸的侵蝕作用將泥沙掏出,而在流速減緩的凸岸,這些泥沙便會沉積下來。這個過程持續不斷,使得邊灘從水下向水上逐漸生長、擴大。邊灘的沉積物顆粒呈現出明顯的分選現象:最靠近主河道、水流較快的地方,沉積的是較粗的礫石和沙;隨著向岸邊抬升,地勢越高,被淹沒的頻率越低,水流能量越弱,沉積物的顆粒也越來越細,最終可能過渡到淤泥和黏土。這種底質梯度,創造了多樣的微棲地。
邊灘的生態意義首先體現在它是河岸植被演替的起點。新生的、裸露的邊灘是惡劣的環境,夏季高溫乾旱,洪水期則被完全淹沒。只有少數耐受性極強的「先驅植物」(pioneer species),如禾本科的草類或某些楊柳科的灌木,能夠在此立足。它們的根系有助於固定不穩定的沉積物,其死亡的殘體則能增加土壤的有機質。隨著時間的推移,土壤條件改善,更多的植物種類得以入侵,逐漸形成草灘、灌木叢,最終可能演化為河岸森林。這個動態的演替過程,為不同階段的昆蟲和動物提供了棲地。例如,裸露的沙灘是某些虎甲蟲和地面蜘蛛的家園,也是燕鷗、鴴科鳥類等灘地築巢鳥類的繁殖地,它們會直接在沙地上產卵,利用沙礫的顏色進行偽裝。
其次,邊灘是水陸生物交換的重要界面。在水位下降的過程中,邊灘上會形成許多孤立的小水窪和潮濕的沙地。這些臨時性的小水體,是蝌蚪、昆蟲幼蟲等小型水生生物的臨時庇護所,也吸引了許多涉禽前來覓食。潮濕的沙地則是許多水生昆-蟲(如蜉蝣、石蠅)羽化成蟲時爬出水面的登陸平台。反之,許多陸生動物,如鹿、野豬,也會來到邊灘飲水或尋找被洪水沖上岸的食物。
河心洲,又稱河道沙洲或礫石灘,是形成於河道中央的沉積體。它通常出現在較寬闊、水流分岔的網狀河(braided river)中,或者在河道突然變寬、流速驟降的區域。與邊灘類似,河心洲也是由水流搬運的沉積物堆積而成,其組成可以是礫石、沙或兩者的混合。由於地處河心,它受到水流的雙面夾擊,形態變化更為劇烈和頻繁。
河心洲最重要的生態功能之一,是為地面築巢鳥類提供了相對安全的繁殖場所。由於四面環水,河心洲能夠有效地將鳥巢與來自陸地的哺乳動物捕食者(如狐狸、鼬、蛇)隔離開來。許多對干擾敏感的鳥類,如燕鷗類、東方環頸鴴等,極度依賴這些河心洲作為其唯一的繁殖地。它們的繁殖成功與否,直接取決於枯水期河心洲的出露面積以及洪水是否來得太早。
此外,河心洲也極大地增加了河道的棲地複雜性。它將原本單一的寬闊河道,分隔成多個流速、深度各不相同的子河道,創造出更多的邊緣棲地。洲的迎水面(上游側)水流較急,類似急瀨;背水面(下游側)則形成一個靜水區,有利於細顆粒沉積和有機物聚集,成為許多小型魚類和無脊椎動物的覓食區和庇護所。河心洲的存在,使得整個河段的棲地類型和水力條件變得更加多樣化,從而能夠支持更豐富的生物群落。
然而,邊灘和河心洲都是極其動態且脆弱的地貌。它們的面積、形態和位置會隨著每一次洪水而改變。這種不穩定性正是其生態價值的一部分,它維持了生態系統的早期演替階段,阻止了整個河岸帶被單一的森林群落完全佔據。不幸的是,現代河流管理常常對這種動態性構成威脅。上游水庫的修建,會攔截大量的泥沙,導致下游的邊灘和河心洲因為得不到泥沙補充而逐漸被侵蝕、消失,這種現象稱為「河床飢餓」(channel starvation)。河道的渠化和堤防的修建,則會固定河岸,阻止河流的自然擺動,使得邊灘無法再生。這些人為干預,都在不知不覺中抹去了河流中這些充滿生機的「沙洲綠洲」,導致依賴它們的生物失去家園,河流生態系統的韌性和多樣性也隨之降低。保護河流的自然輸沙過程和河道擺動的自由,就是保護這些珍貴的灘地生態系。
2.2 底質多樣性:岩盤、礫石、沙與泥的生態意義
河床底質是河流生命的基石,是絕大多數水生生物安身立命的場所。其物理特性——如粒徑大小、穩定性、孔隙度——以及化學特性——如表面積、有機質含量——共同決定了何種類型的生物群落能夠在此繁衍。從上游堅硬的岩盤,到中游鬆散的礫石,再到下游柔軟的泥沙,底質的多樣性直接對應著生物群落結構和功能的多樣性。理解不同底質的生態意義,就如同解讀不同土壤如何孕育不同森林一樣,是通往河流生態系統內部世界的鑰匙。
2.2.1 岩盤環境的固著群落
在河流的某些區段,特別是那些坡度極陡、水流侵蝕力極強的上游峽谷,或是河床由堅硬抗蝕的母岩構成的地方,河床底質可能並非由鬆散的顆粒物組成,而是一整塊連續的、巨大的岩盤(Bedrock)。這種環境在物理上呈現出極端的穩定性和平滑性,為一類特殊的生物群落——固著生物群落(Sessile Communities)——提供了獨一無二的生存舞台。這些生物演化出了非凡的附著能力,將自己牢牢地固定在岩石表面,以抵抗強勁水流的持續衝擊。
岩盤環境最顯著的物理特徵是其極高的穩定性和極低的孔隙度。與礫石或沙質底床不同,岩盤在任何洪水衝擊下都幾乎紋絲不動,這為需要永久性附著點的生物提供了最可靠的基礎。然而,岩盤表面相對平滑,缺乏礫石床那樣複雜的縫隙和避難所。這意味著生活在岩盤上的生物,必須將自己的身體完全暴露在水流之中,直接承受水流的剪應力(shear stress)和攜帶顆粒的磨蝕作用。因此,這裡的生物要麼擁有極其扁平、流線型的體型以減小水流阻力,要麼發展出超強的固著機制。
在這種環境下,**生物膜(Biofilm)**構成了食物網的最底層。在有光照的區域,岩盤表面會被一層由矽藻、綠藻、藍綠藻等微小藻類,以及細菌、真菌和它們分泌的胞外聚合物(Extracellular Polymeric Substances, EPS)混合而成的滑膩薄膜所覆蓋。這層生物膜是岩盤生態系的主要初級生產者,也是許多刮食性動物的直接食物來源。EPS這種黏性物質,是生物膜能夠緊密附著在光滑岩石上的關鍵。
在生物膜之上,更大型的固著生物開始展現其驚人的適應策略。水生苔蘚(Aquatic Mosses)和某些大型絲狀藻類是其中的佼-佼者。它們演化出類似根的「假根」(rhizoids),能夠分泌黏性物質,並鑽入岩石表面微小的裂隙中,將自身牢牢地錨定。它們的群體一旦形成,又能進一步改變局部的微水流環境,為其他更小的生物提供庇護和附著點,形成一個小型的生態系統。
而動物界的固著冠軍,則非黑蠅(Black fly)的幼蟲莫屬。黑蠅幼蟲,又稱蚋,是急流岩盤上最常見的居民之一。它的身體末端有一個特化的「吸盤」,周圍環繞著一圈小鉤,能夠緊緊地鉤在自己事先分泌的一片絲質墊上。這使得它即使在瀑布般的水流中也能穩如泰山。它以一種極為高效的方式濾食:頭部伸出一對精緻的扇狀結構,像捕蝶網一樣在水中展開,攔截漂流而來的細小有機顆粒和藻類。當一個區域的黑蠅幼蟲密度極高時,它們會像一片黑色的地毯一樣覆蓋在岩石上,成為河流生態系中一個強大的生物過濾器。
另一類適應岩盤環境的代表是網石蠶(Net-spinning caddisflies)。這類石蠶蛾的幼蟲不像它們的親戚那樣背著由碎石或植物構成的巢,而是利用從口中吐出的絲,在岩石表面的凹陷處或縫隙中,編織出一個精巧的漏斗狀或帳篷狀的絲網。它們自己則躲在絲網的後方,利用絲網來過濾水流中的食物顆粒。這些絲網的網格大小各不相同,可以捕捉不同尺寸的食物,體現了物種間的資源劃分。
對於魚類而言,光滑的岩盤本身難以提供棲息地,但它們會利用岩盤環境中的其他生物。例如,許多刮食性的魚類(如某些鮎形目魚類)擁有特化的吸盤狀口器,可以吸附在岩石上,穩定身體,同時刮食表面的生物膜。
總而言之,岩盤環境雖然在物理結構上看似單調,但它卻催生了一系列高度特化的生命形態。這裡的生物群落,是一個關於固著、抵抗與過濾的生存故事。它們將堅硬無機的岩石,轉化為一個充滿生命、不斷進行能量轉換的動態表面。在河流生態評估中,這些固著生物的存在與否,常常被用來指示水流的穩定性和底床的受干擾程度。一個被細沙或淤泥覆蓋的岩盤,往往意味著上游存在著嚴重的土壤侵蝕問題,這會導致整個固著生物群落的窒息和崩潰。
2.2.2 礫石層的孔隙生態系
當河流從陡峭的峽谷流出,進入坡度稍緩的河谷,河床便由大塊的岩盤和漂石,過渡到了由大小不一的礫石(Gravel)和卵石(Cobble)構成的鬆散層。這個看似普通的礫石河床,其真正的生態奧秘並不僅僅在於石頭的表面,更在於石頭與石頭之間那片廣闊而隱秘的地下世界——孔隙生態系(Interstitial Ecosystem),或稱間隙帶生態系(Hyporheic Zone Ecosystem)。這是一個三維的、充滿了微小通道和洞穴的迷宮,是地表水與地下水交會的模糊地帶,也是無數微小生命和許多大型生物生命早期階段的庇護所和搖籃。
礫石層的孔隙度(Porosity)是其核心生態價值的來源。大小不一的礫石堆疊在一起,無法完全緊密貼合,它們之間留下的空隙,可以佔到底床總體積的20%到40%。這些孔隙被水流充滿,形成了一個複雜的、與地表主河道相連通的地下水流網絡。水流會從地表河床壓力較高的地方(如下降流區)滲入礫石層,在其中緩慢流動一段距離後,再從壓力較低的地方(如上升流區)湧出,回到主河道。這個水交換過程,使得孔隙中的水體能夠保持相對充足的溶氧和較為穩定的溫度,同時也為其中的生物帶來了細小的有機顆粒作為食物。
這個獨特的孔隙生態系,是大量微小無脊椎動物的家園。這裡棲息著許多體型微小的甲殼類(如橈足類、端足類)、水蟎、線蟲和寡毛類蠕蟲。它們的整個生命週期都在這個黑暗的礫石迷宮中完成。對於這些生物來說,孔隙不僅是它們的家,更是躲避地表急流衝擊和魚類等大型捕食者的完美避難所。它們構成了孔隙生態系中一個複雜的食物網,以滲入的有機碎屑和細菌為食,同時它們自身也成為了更大一些的捕食性無脊椎動物的獵物。
更為重要的是,礫石層的孔隙是許多地表大型生物不可或缺的「育嬰室」。許多我們在河床上常見的水生昆-蟲,如蜉蝣、石蠅和石蠶蛾,它們的卵和極早期的幼蟲階段,都是在礫石間的孔隙中度過的。成蟲將卵產在石頭表面或直接產入水中,卵會被水流帶入礫石縫隙中。在這裡,卵和剛孵化的微小幼蟲可以免受水流的直接衝擊和捕食者的威脅,在一個相對穩定和安全的環境中完成其最脆弱的生命階段。直到它們長得足夠大,具備了抵抗水流和躲避敵害的能力後,才會逐漸遷移到礫石的表面活動。
對於許多魚類來說,礫石層的健康狀況直接關係到它們的繁殖成敗。最具代表性的就是鮭科魚類(如鮭魚、鱒魚)。在繁殖季節,雌魚會用尾巴奮力地扇動河床,在精心挑選的礫石區挖掘出一個稱為「產卵巢」(redd)的凹坑。她將卵產在坑中,雄魚隨即為其授精,然後雌魚會到凹坑的上游,再次扇動礫石,讓水流將較小的礫石沖下,輕輕地覆蓋在魚卵之上。這個礫石「被子」的厚度和孔隙度至關重要。它必須足夠厚,以保護魚卵不被水流沖走或被其他動物吃掉;但同時,它的孔隙又必須足夠通暢,以保證有充足的、富含氧氣的水流流過魚卵,供給它們呼吸所需,並帶走代謝產生的廢物。如果礫石層的孔隙被過量的細沙或淤泥堵塞(這個過程稱為淤積作用 Siltation),流經魚卵的水流就會中斷,導致魚卵因缺氧而大批死亡。因此,一個乾淨、疏鬆、孔隙度高的礫石河床,是維持這些魚類族群健康的生命線。
總結來說,礫石層的生態意義遠超其表面所見。它所創造的孔隙生態系,是一個隱藏的、三維的生物多樣性熱點。它不僅為大量特化的微型生物提供了永久的家園,更為河流中許多重要物種(包括具有重要經濟和生態價值的魚類)的生命早期階段提供了不可替代的庇護和孵化場所。然而,這個脆弱的地下世界正受到人類活動的嚴重威脅。來自上游農業、林業、道路建設等活動所造成的土壤侵蝕,會帶來大量的細顆粒泥沙,像水泥一樣堵塞礫石層的孔隙,導致整個孔隙生態系的窒息和崩潰。保護礫石河床的清潔和完整,是河流保育工作中一項至關重要但又常常被忽視的任務。
2.2.3 沙質底床的生物活動
當河流的坡度進一步減緩,水流的能量已不足以搬運礫石時,沙(Sand)便成為了河床底質的主角。沙質底床廣泛分布於中游的緩流區、邊灘以及下游河段。與岩盤的穩定和礫石的粗糙相比,沙質底床呈現出截然不同的物理特性:高度的不穩定性和均質的顆粒。沙粒細小(粒徑介於0.0625至2毫米之間),在水流作用下極易移動,形成沙波(sand ripples)或沙丘(dunes),整個河床形態處於一種持續的、動態的變化之中。這種不穩定的環境,對於需要固著基質的生物來說是個災難,但它也催生了一套獨特的、以鑽穴(burrowing)和快速移動為核心的生存策略。
生活在沙質底床的生物,首先必須解決的問題就是如何在一個不斷移動的基質中保持自己的位置。因此,這裡的生物群落幾乎完全由能夠快速鑽入沙中,或在沙中自由活動的物種組成。它們將沙層本身作為躲避水流衝擊和天敵捕食的庇護所。許多物種演化出了適應鑽沙的特殊形態。例如,某些蜉蝣的幼蟲身體呈魚雷狀,頭部扁平如鏟,足部強壯有力,能夠在幾秒鐘內將自己埋入沙中。一些雙殼類的軟體動物(如蚌、蛤)擁有斧狀的足,可以像船錨一樣插入沙中,將身體固定。
沙質底床的孔隙空間遠小於礫石層,且容易被更細的顆粒堵塞,因此其內部的氧氣交換較差。然而,在表層幾公分的沙層中,由於水流的擾動和生物自身的活動,依然可以維持一定的溶氧水平。這裡的生物群落主要由「採集性集食者」(gathering-collectors)主導。它們的食物來源是沉降在沙粒之間或沙表面的細顆粒有機物質(FPOM)、細菌和微藻。典型的代表是大量的搖蚊(Chironomidae)幼蟲和寡毛類蠕蟲。它們在沙層中建造臨時性的管道或直接在顆粒間穿行,吞食富含有機質的沉積物。
**生物擾動(Bioturbation)**是沙質底床生態系中一個極其重要的過程。這些鑽穴動物在沙中不斷地移動、覓食和建造居所,它們的活動就像無數微小的犁,持續地翻動和混合著表層的沉積物。這種擾動產生了多方面的生態效應。首先,它增加了沙層的孔隙度和透水性,促進了氧氣和營養物質向更深層的滲透,擴大了宜居的空間。其次,它將深層的、已被微生物分解的有機物帶到表層,同時將表層新鮮的有機物帶入深層,加速了營養物質的再循環過程。最後,生物擾動本身也改變了底床的微地形,創造了更多樣的微棲地。可以說,這些不起眼的鑽穴生物,是沙質底床生態系的「生態工程師」(ecosystem engineers)。
對於魚類而言,沙質底床提供了獨特的覓食機會。許多底棲魚類,如鰍科魚類,擁有特化的、朝向下方的口和靈敏的觸鬚,它們會貼著河床遊動,不斷地用吻部探測和翻攪沙子,尋找藏在其中的無脊椎動物。另一些魚類,如比目魚或鰈魚,則演化出了扁平的身體和隨環境改變體色的能力,它們會將自己半埋在沙中,伏擊游過的小魚或甲殼類,沙質底床為它們提供了完美的偽裝。
然而,沙質底床的不穩定性也帶來了挑戰。一場較大的洪水,就可能將整個沙床完全重塑,將原有的生物群落沖走或深埋。因此,這裡的生物通常具有很強的恢復力(resilience)。它們往往具有較短的生命週期、高繁殖率和強大的擴散能力,能夠在災難性的干擾過後,迅速地從周邊未受影響的區域重新移入,並建立新的族群。這種「機會主義」(opportunistic)的生活史策略,是對動盪環境的完美適應。
總結來說,沙質底床是一個充滿動感和變化的環境。它篩選出了一群以鑽穴為生的「沙中居民」,它們的生物活動反過來又深刻地影響著底床的物理和化學特性。這個生態系統雖然在物種豐富度上可能不如結構複雜的礫石河床,但其在物質循環和對干擾的快速響應方面,展現了獨特的生態功能。在進行河道工程時,需要認識到沙的自然流動和堆積是河流健康的一部分,過度挖沙或固定河床,都會破壞這個以動態為本質的生態系統。
2.2.4 泥質環境的有機物累積
當河流的旅程接近尾聲,進入廣闊的下游平原、河口或流速極緩的迴水區時,水流的能量已極度衰減,連最輕、最細的顆粒——淤泥(Silt)和黏土(Clay)——也得以沉降下來。這些細微的顆粒與大量的有機碎屑混合,形成了厚厚一層柔軟、黏稠的泥質底床。這個環境在物理和化學特性上與上、中游的底質有著天壤之別,它是一個巨大的有機物倉庫和生物化學反應器,孕育了一套專門適應低氧、高有機質環境的生物群落。
泥質環境最核心的特徵是極低的孔隙度和極高的有機質含量。淤泥和黏土的顆粒極其微小,它們緊密地堆積在一起,幾乎沒有任何孔隙可言。這導致水體在泥層內部的交換極為緩慢,氧氣很難滲透到表層以下。與此同時,從整個流域匯集而來的細顆粒有機物質(FPOM)在此大量沉積,與泥沙混合在一起。大量的微生物(主要是細菌)會分解這些有機物,而在這個過程中會消耗掉本已稀少的氧氣。因此,泥質底床的表層以下幾毫米,通常就已經是**缺氧(hypoxic)甚至無氧(anoxic)**的狀態。在無氧條件下,厭氧細菌會通過不同的化學路徑(如硫酸鹽還原)來分解有機物,這個過程會產生硫化氫(H₂S)等有毒氣體,這也是許多池塘或河口底泥會散發出臭雞蛋氣味的原因。
在這樣一個看似惡劣的環境中,生命依然找到了出路。這裡的生物群落幾乎完全由能夠耐受低氧和高有機質的沉積物攝食性集食者(deposit-feeding collectors)所主導。它們直接吞食富含有機質的底泥,消化其中的細菌、藻類碎屑和有機質,然後將無機的礦物顆粒排出。最具代表性的生物就是顫蚓科(Tubificidae)的蠕蟲,俗稱「水蚯蚓」。它們通常將身體的前半部分鑽入缺氧的泥中覓食,而後半部分則伸出到泥水界面,暴露在相對富氧的水中。它們的尾部富含微血管,皮膚極薄,可以作為呼吸器官,在水中不停地擺動以增加氣體交換。更神奇的是,顫蚓的血液中含有高濃度的血紅蛋白(hemoglobin),使其呈現出鮮紅色。這種血紅蛋白對氧氣有極高的親和力,使得它們能夠在極低溶氧的環境下,依然能高效地捕獲和運輸氧氣。因此,顫蚓的大量出現,常常被視為水體受到嚴重有機污染的指標。
另一類泥質環境的優勢物種是**搖蚊科(Chironomidae)**的某些幼蟲。與顫蚓類似,許多生活在底泥中的搖蚊幼蟲(常被稱為紅蟲)也含有血紅蛋白,使它們能夠在低氧環境下生存。它們會在泥中建造U型或直立的管狀居所,並通過身體的節律性擺動,在管中產生一股微弱的水流,將新鮮的、含氧的水帶入管中,同時也帶來了食物顆粒。
除了這些小型無脊椎動物,一些大型的**雙殼貝類(Bivalvia)**也適應了泥質環境。它們將斧足插入泥中固定身體,大部分軀體埋在泥下以躲避捕食者,只伸出兩根長長的水管(siphon)到泥層之上的水中。一根是入水管,將水和其中的懸浮食物顆粒(浮游植物和有機碎屑)吸入體內進行過濾;另一根是出水管,將過濾後的水和排泄物排出。它們是泥質環境中強大的生物過濾器。
泥質環境的有機物累積和分解功能,使其在整個河流生態系統的物質循環中扮演著終端處理者的角色。這裡是碳、氮、磷等元素最終的匯集地(sink)。微生物的分解作用,將複雜的有機物轉化為無機的營養鹽。這些營養鹽一部分會被重新釋放到水體中,供浮游植物利用,啟動新的生產循環;另一部分則可能被永久地埋藏在沉積物中。這個過程的速率和效率,對下游水域乃至河口和近海的生態健康有著深遠的影響。
總結而言,泥質環境是一個由有機物驅動、以分解作用為核心的生態系統。它雖然物種多樣性相對較低,但優勢物種的生物量卻可能極其龐大。這裡的生物展現了對低氧和化學逆境的極致生理適應。理解泥質環境的生物地球化學過程,對於評估河流的自淨能力、管理有機污染以及預測下游優養化的風險至關重要。
2.3 水流的塑造力:侵蝕、搬運與堆積作用
水流,是河流的靈魂,也是最強大的地貌雕塑家。一條河流的樣貌,從宏偉的峽谷到廣闊的平原,從微小的沙波到巨大的沙洲,無一不是水流力量的傑作。這個塑造過程主要通過三種緊密相連的作用來完成:侵蝕(Erosion)、搬運(Transportation)和堆積(Deposition)。這三者之間的平衡與轉換,由水流的能量(主要取決於流速和流量)所決定。當水流能量足以克服顆粒的重力和摩擦力時,侵蝕和搬運便會發生;當能量衰減到無法再承載其負荷時,堆積便隨之而來。這個永不停歇的過程,不僅僅是地理學上的變遷,它更深刻地影響著棲地的穩定性、資源的可利用性,以及生物群落的結構與演替。
2.3.1 侵蝕作用對棲地的影響
侵蝕作用是水流塑造力的起點,是河流從其河床和兩岸獲取建築材料(沉積物)的過程。它主要發生在水流能量最強勁的區域,如上游的陡峭河段、曲流的凹岸以及瀑布的下方。侵蝕不僅僅是簡單的沖刷,它通過多種方式對棲地產生深刻而持久的影響,既有創造性的一面,也有破壞性的一面。
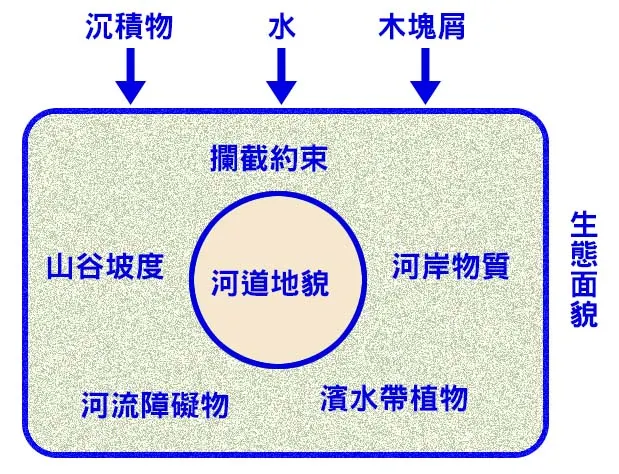
侵蝕的創造性體現在它能夠開拓和加深棲地空間,並增加物理結構的複雜性。在堅硬的基岩河床上,水流攜帶著沙礫等磨料,像砂紙一樣對河床進行持續的「磨蝕作用」(abrasion),能夠在岩石上刻劃出溝槽、坑洞,甚至形成壯觀的壺穴(potholes)。這些微地形為小型生物提供了寶貴的庇護所。更為宏觀的侵蝕,則造就了河流最基本的形態。例如,在曲流凹岸的持續性側向侵蝕,不僅加深了水體,形成了為大型魚類所偏愛的深潭棲地,同時也可能掏空岸邊的樹根,導致樹木倒塌入水。這些倒木(large woody debris)是極其重要的棲地結構,能夠創造出複雜的渦流和庇護空間。河流的下切侵蝕,則在漫長的地質年代中,切割出深邃的峽谷,創造了獨特的、與世隔絕的生態環境,有時甚至會促進新物種的形成。
然而,侵蝕也具有強烈的破壞性,特別是在其速率因自然或人為因素而加劇時。一場極端的洪水,其巨大的侵蝕力可以在短時間內徹底改變一個河段的面貌。它可以將原本穩定的礫石灘完全沖毀,填平深潭,導致棲地均質化。這種災難性的物理干擾,會對原有的生物群落造成毀滅性打擊,將其「重置」到一個非常早期的演替階段。
更為常見的是由人類活動引發的加速侵蝕。當流域內的森林被砍伐、土地被過度開墾或進行大規模工程建設時,裸露的土壤失去了植被的保護,在降雨時會被大量沖刷入河。這導致河流中的懸浮沉積物濃度急劇升高。高濃度的懸-浮沙礫,會像噴砂機一樣,對河床中的生物進行強烈的物理磨損。它們會刮走附著在石頭上的藻類和生物膜,直接傷害或殺死固著的無脊椎動物,並磨損魚類的鰓和體表,增加其感染疾病的風險。這種增強的磨蝕效應,會極大地降低河流的初級生產力和生物多樣性。
此外,不當的河道管理也會引發意想不到的侵蝕問題。例如,在河道中過度採砂,會使得河床局部下降,形成一個「侵蝕基準面」,這會引發向上游和下游的「溯源侵蝕」(headward erosion)和「向源侵蝕」,導致橋墩裸露、堤防根基被掏空等工程安全問題。同時,河床的普遍下切,也會導致地下水位下降,影響到周邊的農業和植被。
總結來說,侵蝕是河流一種天然且必要的動態過程,它在自然的節奏下,是創造棲地多樣性的重要力量。然而,當侵蝕的速率和規模超出自然變動的範圍時,它就會變成一種強大的破壞力,不僅威脅到水生生物的生存,也可能對人類社會的基礎設施構成風險。因此,流域的土地利用管理,特別是水土保持工作,是控制河流侵-蝕速率、維護健康棲地的根本所在。
2.3.2 搬運過程的生態動態
一旦沉積物被水流侵蝕剝離,它便進入了生命週期中的第二個階段——搬運(Transportation)。搬運是水流將其攜帶的泥沙、礫石等物質從一個地方移動到另一個地方的過程。這個過程不僅僅是物質的位移,它本身就是一個充滿動態變化的生態過程,深刻影響著水體的物理化學性質、生物的行為與分布,以及能量在河流系統中的傳遞。
沉積物的搬運方式取決於顆粒的大小和水流的能量。最細微的黏土和淤泥顆粒,可以長時間地懸浮在水體中,隨著主流一起運動,這被稱為懸浮載(Suspended Load)。它們是造成中、下游河水混濁的主要原因。懸浮載的濃度,直接影響光的穿透深度,從而控制著水體中光合作用的強度。高濃度的懸浮載會抑制浮游植物和底棲藻類的生長,降低河流的初級生產力。同時,這些懸浮顆粒也會對濾食性生物造成影響,過高的無機顆粒濃度可能會堵塞它們的濾食器官,或降低它們獲取食物的效率。
較大的顆粒,如沙和細礫,則以**推移載(Bed Load)的方式在河床底部移動。它們的運動方式多種多樣:較小的沙粒會以跳躍(saltation)的方式,在水流的舉升力作用下,一跳一跳地前進;稍大的顆粒則以滾動(rolling)或滑動(sliding)**的方式,貼著河床移動。推移載的運動,是塑造河床微地貌(如沙波、沙丘)的主要力量。這個持續移動的底床,對於生物來說是一個極具挑戰性的環境。任何不能快速鑽穴或及時躲避的底棲生物,都可能被移動的沙礫磨損、掩埋或沖走。因此,推移載活躍的河段,其底棲生物的密度通常較低,且物種組成也偏向於那些具有高度移動性和快速繁殖能力的機會主義者。
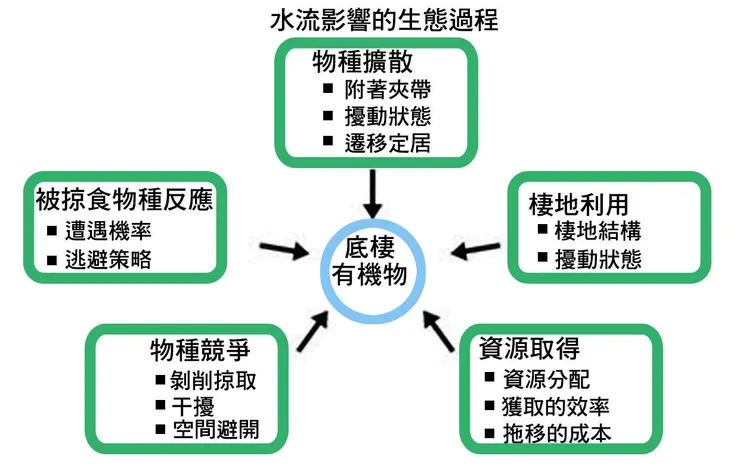
推移載的運動,也催生了一種獨特的生態現象——漂流(Drift)。生活在河床礫石間的許多水生昆-蟲幼蟲,有時會主動或被動地進入水體,隨著水流向下游漂移一段距離。這種行為的成因複雜,可能是為了尋找新的食物資源、躲避捕食者、逃離過於擁擠的棲地,或是對環境壓力(如水溫變化、化學污染)的反應。無論原因如何,這種生物性的搬運過程,是無脊椎動物在河流中進行種群擴散和基因交流的重要機制。然而,漂流也伴隨著巨大的風險,因為漂浮在水中的幼蟲極易被魚類捕食。事實上,這種「無脊椎動物漂流」,構成了許多河流魚類最主要的食物來源。魚類會佔據有利位置,捕食這些順流而下的「外賣」,形成了河流生態系中一條關鍵的能量傳遞鏈路。
從更宏觀的尺度看,沉積物的搬運過程是連接整個流域的紐帶。上游侵蝕的礫石,可能在中游的邊灘上短暫停留,然後在下一次洪水中繼續它的旅程,最終可能以細沙的形式到達河口。在這個過程中,顆粒物本身也在不斷地被磨損、分選。這種物質的連續輸送,確保了下游生態系統能夠獲得來自上游的礦物質和部分有機質供給。然而,人類活動對這個搬運過程造成了巨大的干擾。最典型的例子就是水庫的修建。大壩如同一道巨大的屏障,攔截了幾乎所有的推移載和大部分的懸浮載,使其在水庫中沉積下來。這導致大壩下游的河流處於一種「飢餓」狀態,因為失去了正常的沉積物補給,水流的能量會轉而侵蝕自身的河床和河岸,造成河床下切、棲地破壞等一系列問題。
總而言之,搬運過程是河流動態本質的體現。它不僅塑造地貌,影響水質,更驅動著生物的行為和物種間的相互作用。理解沉積物如何被搬運,以及這個過程如何被干擾,對於我們評估河流健康、管理漁業資源以及制定可持續的基礎設施建設計劃都至關重要。
2.3.3 堆積環境的生物群落
當水流的能量減弱到無法再支撐其攜帶的沉積物負荷時,搬運過程便宣告結束,取而代之的是堆積(Deposition)或沉積(Sedimentation)作用。堆積是水流塑造力的最終章,它將從上游攜來的物質財富,鋪設在河床之上,創造出全新的陸地和水下地貌。這些堆積環境,如中游的邊灘、下游的沖積平原和河口三角洲,雖然在物理上看似平緩單調,卻是河流生態系中生產力最高、生物化學過程最活躍的區域之一,支持著獨特而豐富的生物群落。
堆積環境的形成,與水流能量的耗散直接相關。這通常發生在幾個關鍵區域:在曲流的凸岸,水流速度減慢,導致沙礫堆積成邊灘;在河道突然變寬或匯入湖泊、水庫的地方,水流擴散,能量驟降,會形成水下三角洲;而在河流入海口,淡水與密度更高的海水相遇,流速銳減,加上鹽水中的離子會促使懸浮的黏土顆-粒絮凝成更大的團塊,加速沉降,形成了廣闊的三角洲和潮灘。這些環境的共同特點是,底質主要由較細的顆粒(沙、淤泥、黏土)組成,且富含有機質。
在這些以細顆粒和有機質為特徵的堆積環境中,生物群落的結構發生了根本性的轉變。由於缺乏穩定的硬質基底,固著生物和刮食性生物幾乎完全消失。取而代之的,是一個由**沉積物攝食者(Deposit Feeders)和濾食者(Filter Feeders)**主導的群落。它們演化出各種巧妙的策略,來利用這個富饒但鬆軟的環境。
沉積物攝食者是堆積環境中最具代表性的功能群。它們直接吞食富含有機質的底泥,消化其中的微生物和有機碎屑。這個群體包括了大量的寡毛類蠕蟲(如顫蚓)、多毛類蠕蟲(在河口鹹水區)以及多種搖蚊幼蟲。它們在柔軟的底泥中鑽穴而居,其活動(生物擾動)深刻地改變著沉積物的物理和化學性質,促進了氧氣的滲透和營養鹽的釋放,是堆積環境中名副其實的「生態工程師」。
濾食者則利用另一種策略。它們將身體固定在沉積物中,通過特化的器官過濾流經水體中的懸浮食物顆-粒,主要是浮游植物和細顆粒有機物。大量的雙殼貝類(如蚌、蛤、牡蠣)是這個功能群的代表。它們在河口和下游的淺灘區,常常能形成密度極高的群落(如牡蠣礁),其強大的濾水能力,能夠顯著提高水體的透明度,對控制藻華、維持水質具有至關重要的作用。它們堅硬的殼體在死後會留在原地,又為其他生物(如藤壺、海葵、小型魚類)提供了寶貴的硬質附著基底,進一步增加了棲地的複雜性。
堆積環境,特別是河口和洪氾平原,是極其重要的育幼場(Nursery Grounds)。週期性的洪水將營養物質帶到洪氾平原上,刺激了植物和浮游生物的爆發性生長。許多魚類和甲殼類(如蝦、蟹)會利用這個時機,進入這些被淹沒的淺水區產卵和覓食。這裡豐富的食物和複雜的植被結構,為脆弱的幼體提供了充足的營養和有效的庇護,使其能夠快速成長。可以說,許多具有重要商業價值的漁業資源,其產量的豐歉,直接取決於這些堆積環境的健康和完整性。
此外,露出水面的堆積體,如沙洲和泥灘,是遷徙水鳥賴以生存的驛站。在漫長的遷徙旅途中,數以百萬計的鴴鷸類水鳥,需要在這些灘地上停歇、覓食,補充能量。灘地中豐富的底棲無脊椎動物(如蠕蟲、貝類),是它們主要的食物來源。這些堆積環境的面積和食物豐度,直接關係到遷徙鳥類能否成功完成其生命史中最艱鉅的旅程。
總結來說,堆積環境是河流將其從流域各處收集的物質和能量進行整合、再分配和再利用的場所。它雖然缺乏上游棲地的物理多樣性,但其巨大的生物量、高效的物質循環和對關鍵物種生命史的支撐作用,使其在整個生態系統中具有不可替代的價值。保護這些堆積環境,意味著保護河流的自淨能力、漁業的未來以及全球尺度的生物遷徙廊道。
2.3.4 河道地貌的演化過程
河流的地貌並非一成不變,而是在水流的持續作用下,經歷著一個從短期(單次洪水)到長期(千百年地質年代)不斷演化的動態過程。這個演化過程,是河流對其所在流域的氣候、地質、地形和植被等條件進行響應和自我調節的結果。理解河道地貌的演化,能讓我們跳出靜態的視角,將河流視為一個有生命、會成長、會變老的動態實體,從而更深刻地把握其生態系統的變遷規律。
河道地貌的演化,可以從不同的空間形態上來理解,主要包括河道的平面形態、縱剖面和橫剖面的變化。
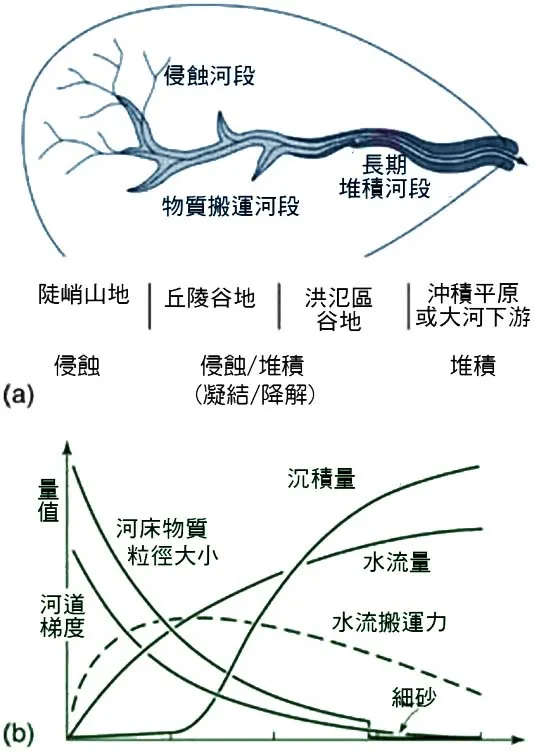
平面形態的演化,指的是河道在俯瞰視角下的形狀變化,最主要的三種類型是直線型(Straight)、曲流型(Meandering)和網狀型(Braided)。很少有河流是天然筆直的,即使在看似直線的河段,其最深線(thalweg)也往往是彎曲的。更常見的是曲流型河道,它主要發育在坡度較緩、河岸由較細沉積物構成的中下游平原。曲流的演化是一個自我加強的過程:一個微小的彎曲,就會引發凹岸侵蝕和凸岸堆積,使得彎曲的幅度越來越大。隨著時間的推移,曲流的頸部會變得越來越窄,最終在一次大洪水中,河流可能會「截彎取直」(neck cutoff),廢棄原有的彎曲河道,形成一個月牙形的湖泊——牛軛湖(Oxbow Lake)。牛軛湖是洪氾平原上極其重要的濕地生態系統,它會隨著時間逐漸淤積,經歷從開闊水域到沼澤,再到陸地森林的完整生態演替過程。
網狀河道則發育在那些坡度較陡、沉積物供給非常豐富(特別是粗顆粒的推移載)的地區,如山麓沖積扇或冰川出水口。在這些地方,水流攜帶著超出其搬運能力的沉積物,導致沉積物在河道中不斷堆積,形成大量的河心洲。這些沙洲會迫使水流分岔、匯合、再分岔,形成一個像辮子一樣交織的、寬闊而淺的河道網絡。網狀河道的形態極不穩定,每一次洪水都可能導致舊的沙洲被沖毀,新的沙洲在別處形成,主水道的位置也隨之改變。這種高度動態的環境,為那些適應頻繁干擾的先驅物種提供了廣闊的舞台。
縱剖面的演化,指的是沿著河流從源頭到河口的坡度變化。一條理想化的成熟河流,其縱剖面是一條平滑的、上陡下緩的凹形曲線,這被稱為「平衡剖面」(profile of equilibrium)。河流會通過侵蝕和堆積作用,不斷地調整其縱剖面,以達到一種狀態:在這種狀態下,水流的能量剛好足以搬運其所能獲得的沉積物,既不產生淨侵蝕,也不產生淨堆積。任何對這個平衡的干擾,都會引發河流的自我調整。例如,如果海平面下降(或陸地抬升),河口的侵蝕基準面降低,河流就會從下游開始向上游進行溯源侵蝕,以適應新的基準面,這個過程會在地貌上留下一系列的「河流階地」(river terraces),記錄了河流過去的演化歷史。
橫剖面的演化,則反映了河道寬度和深度的變化。在上游,河流能量主要用於下切侵蝕,形成深而窄的「V」形谷。到了中下游,側向侵蝕變得更加重要,河道拓寬,形成了寬闊的河谷和洪氾平原。河道的橫剖面形狀,也與河岸的物質組成和植被狀況密切相關。植被茂密的河岸更能抵抗侵蝕,有助於維持一個較窄、較深的河道;而被破壞的河岸則容易崩塌,導致河道變寬、變淺。
人類活動正在以前所未有的速度和規模干擾著河道地貌的自然演化。水庫建設、河道渠化、城市化導致的不透水面增加、土地利用變化導致的輸沙量改變等等,都在迫使河流偏離其自然的演化軌跡。例如,渠化將蜿蜒的曲流變成了筆直的排水溝,雖然提高了水流速度,卻徹底摧毀了急流-深潭序列和多樣的棲地,切斷了河流與洪氾平原的聯繫。理解河道地貌的自然演化規律,是進行科學的河流管理和生態修復的基礎。成功的河流修復,往往不是要將河流固定在某一個理想的形態,而是要恢復其自然的水文和輸沙情勢,給予河流足夠的空間和自由,讓它能夠重新啟動其自我調節和演化的能力,從而創造出一個動態、健康且富有韌性的生態系統。
2.4 季節流量變化:洪水與枯水期的生態衝擊
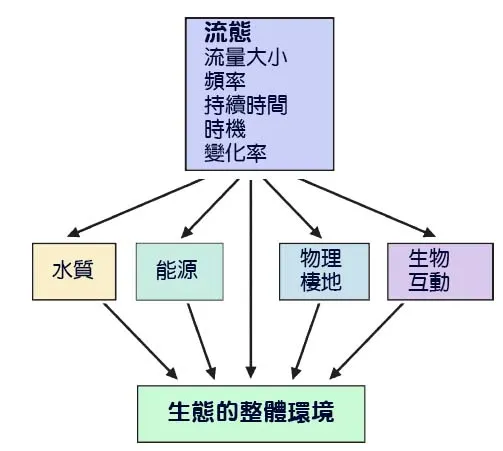
如果說河道地貌是河流的骨架,那麼流量(Discharge)——單位時間內通過河道某一斷面的水量——就是流淌其中的血液。河流的流量並非恆定不變,而是隨著季節性的降雨、融雪和蒸發而發生劇烈的波動,形成週期性的洪水期(High Flow)和枯水期(Low Flow)。這種流量的變化,被稱為「水文情勢」(Flow Regime),是河流生態系統最核心的驅動變量之一。洪水和枯水,並非單純的水量增減,它們是兩種截然不同的生態力量,從根本上重塑棲地、篩選生物、並調節著整個生態系統的能量流動和生命週期。理解洪水與枯水期的生態衝擊,是解讀河流生命如何適應並利用環境動態性的關鍵。
2.4.1 洪水期的棲地重塑
洪水,常常在人類社會中被視為具有破壞性的天災。然而,從生態學的角度來看,週期性的、在自然變動範圍內的洪水,是河流生態系統進行新陳代謝、維持健康與活力的必要過程。它是一股強大的重塑力量,通過物理和化學的雙重作用,對河流棲地進行一次大規模的「大掃除」和「再創造」,其影響深遠而複雜。
洪水期最直接的物理衝擊,來自於流量和流速的劇增。當巨大的水量湧入河道,水流的能量呈指數級增長。這股強大的力量,足以移動河床上那些在平水期看似紋絲不動的巨石,將淤積的沙洲夷為平地,或是在新的地方創造出沙洲。這種對河床底質的劇烈攪動和重置,被稱為「底床擾動」(substrate mobilization)。這個過程對底棲生物群落來說,無疑是一場浩劫。許多固著或行動緩慢的生物會被直接沖走或被移動的石塊砸傷、掩埋。然而,這種看似毀滅性的過程,卻有著至關重要的生態功能。它能夠沖刷掉堵塞在礫石層孔隙中的細沙和淤泥,恢復礫石產卵場的通透性,為下一輪魚類的成功繁殖和無脊椎動物幼蟲的生存清理出「育嬰室」。同時,它也能清除掉過度生長的藻類和水生植物,防止其過度繁盛而導致的物種單一化,為新的物種入侵和群落演替創造出空間。
其次,洪水是重塑河道宏觀地貌的主要動力。只有在洪水期間,河流才有足夠的能量去進行顯著的侵蝕和堆積。洪水會切割曲流的凹岸,擴大深潭的體積;會將大量的沉積物搬運到凸岸,擴建邊灘;甚至可能截彎取直,創造出新的牛軛湖。這種地貌的重塑,維持了「急流-緩流-深潭」序列的動態平衡,防止了河道因長期淤積而變得均質化。可以說,河流的棲地多樣性,是在一次次的洪水中被反覆雕琢和維護的。
第三,洪水是實現橫向連通性的關鍵機制。當水位上漲,漫過常規的河岸,河流便與其廣闊的洪氾平原連接在一起。這不僅僅是水域面積的擴大,更是兩個生態系統之間物質、能量和生物的密集交換。洪水將主河道中的營養鹽和生物(如魚類、浮游生物)帶到洪氾平原,觸發了那裡驚人的生產力爆發。同時,洪水也將陸地上的有機物質(如枯枝落葉、土壤有機質)帶入水體,為水生食物網注入新的能量。這種「洪水脈衝」(flood pulse)是維持大型河流生態系統高生產力的核心驅動-力。
此外,洪水還扮演著**「清道夫」**的角色。在枯水期,河道中可能會累積大量的有機廢物和污染物。一場洪水能夠將這些累積物稀釋並迅速輸送至下游,在一定程度上實現了河流的「自我淨化」。當然,這也可能將污染物擴散到更廣泛的區域。
對於生物而言,洪水是一場嚴峻的考驗,但也是演化適應的驅T-shirt。許多生物演化出了特殊的行為來應對洪水。一些魚類會躲藏在洪水流速較慢的避難所,如岸邊的淹沒植被中、倒木後方或支流匯入口。許多無脊椎動物則會向更深的底床孔隙中鑽去(垂直避難),或者利用其流線型的身體緊貼在石頭的背水面。那些無法有效躲避的物種會被淘汰,而能夠存活下來的,則是對這種干擾具有高度適應性的物種。因此,洪水也是一個強大的自然選擇力量,塑造了河流生物的物種組成和生活史特徵。
總結來說,洪水期是河流的「重啟」和「更新」期。它通過劇烈的物理擾動,打破舊有的平衡,清理廢物,重塑棲地,並重新連接水陸生態系統。一個失去了自然洪水脈衝的河流(例如被大壩完全控制的河流),就像一個失去了發燒和免疫反應能力的身體,會逐漸因為內部廢物的累積、棲地的老化和單一化,而失去其生命的活力與韌性。
2.4.2 枯水期的棲地限制
與洪水期的狂野奔放形成鮮明對比,枯水期(Low Flow)是河流一年中最低調、最內斂的時期。在經歷了季節性降雨的間歇後,河流的流量下降到最低點,水位收縮,許多在平水期位於水下的區域暴露出來。這個時期,水流的塑造力減到最弱,取而代之的是一系列由水量減少引發的物理和化學「壓力」,對河流生物構成了嚴峻的生存考驗。枯水期,是一個棲地被壓縮、競爭加劇、環境質量可能惡化的限制性時期。
枯水期最直接的影響是物理棲地空間的急劇縮減和破碎化。隨著水位下降,寬闊的河道可能會收縮成一條狹窄的水流,原本連續的棲地被分割成一個個孤立或半孤立的水體。急瀨區域可能完全乾涸,只剩下濕潤的石頭;廣闊的緩流區可能變成一系列淺灘和獨立的小水窪。這種棲地的喪失和破碎化,對水生生物產生了巨大的壓力。首先,生物的總承載量下降,所有生物都被迫擠壓在有限的剩餘水體中。其次,棲地類型的多樣性降低,特別是對於那些依賴特定水深或流速的物種,它們可能完全失去賴以生存的家園。
棲地空間的壓縮,不可避免地導致了生物間相互作用的加劇。在擁擠的水體中,種內和種間的競爭變得異常激烈。無論是爭奪有限的食物資源,還是搶占深潭中少數能提供庇護和涼爽水溫的優質領地,競爭的失敗者都可能面臨死亡或被迫遷徙。同時,捕食壓力也顯著增加。由於獵物(如小型魚類和無脊椎動物)的密度增高,且可供躲藏的空間減少,捕食者(如大型魚類、鳥類)的捕食效率會大大提高。對於獵物而言,枯水期是一個極其危險的時期。許多被困在淺水窪中的生物,會成為一場捕食者的盛宴。
除了物理空間的限制,枯水期還常常伴隨著水質的惡化。由於總水量減少,河流對於外來污染物的稀釋和自淨能力會大幅下降。在平水期看似無害的、持續排入的少量污染物(如生活污水、農業廢水),在枯水期會因為水量減少而濃度急劇升高,可能達到對生物產生毒害的水平。此外,緩慢的水流和強烈的日照,會使得淺水區的水溫迅速升高,有時甚至會超過許多生物的耐受上限。高溫還會降低水中的溶氧量,而擁擠的生物群落進行呼吸作用,會進一步消耗氧氣。在極端情況下,尤其是在夜晚,某些孤立的水潭可能會出現嚴重的缺氧現象,導致生物集體死亡。
因此,枯水期成為了一個強大的環境篩選器。只有那些能夠耐受高溫、低氧、高密度和激烈競爭的物種才能存活下來。同時,**避難所(Refugia)**的存在在枯水期變得至關重要。那些在枯水期依然能保持足夠水深和較低水溫的深潭,或者有穩定地下水補給的支流匯入口,就成為了生物的「諾亞方舟」。大量的魚類和其他生物會聚集在這些避難所中,度過最艱難的時期。這些避難所的數量和品質,直接決定了一個河段的生物群落能夠在枯水期過後恢復到何種程度。
總結來說,枯水期是河流生態系統的「瓶頸期」。它通過壓縮棲地、加劇生物互動和惡化水質,對生物施加了強烈的選擇壓力。這個時期雖然充滿挑戰,但它也淘汰了不適應的個體,並凸顯了優質棲地(特別是避難所)的極端重要性。人類活動,如過度取水灌溉或工業用水,會人為地加劇枯水期的嚴重程度和持續時間,對河流生態系統造成毀滅性的打擊。維持河流在枯水期必要的「生態基流量」(environmental flow),是確保河流生命能夠度過難關、得以延續的最低要求。
2.4.3 流量變化的生物適應
面對洪水期毀滅性的物理衝擊和枯水期嚴苛的生存壓力,河流中的生物並非束手無策的受害者。在漫長的演化歷程中,它們發展出了一系列精妙的形態、生理和行為上的適應策略,使其不僅能夠在這種動態變化的環境中存活下來,甚至能夠巧妙地利用流量變化所帶來的機會。這些適應策略,是生命應對環境節律的智慧結晶,也是河流生態系統充滿韌性的根本原因。
在形態適應方面,許多生物演化出特殊的身體結構來應對水流的挑戰。為了抵抗洪水期的急流,生活在急瀨的許多無脊椎動物都具有流線型或扁平的身體,如扁蜉蝣(Heptageniidae),它們的身體像一片緊貼在石頭上的葉子,極大地減小了水流的舉升力。許多魚類,如鱒魚和石𩼧,也擁有完美的紡錘形流線型身體,使其能夠以最小的能量消耗在湍急的水中保持位置或快速移動。為了在洪水中固定自己,黑蠅幼蟲演化出了末端的吸盤,網石蠶則會編織絲網,而許多蜉蝣和石蠅的幼蟲則擁有強壯的、帶有爪子的足,可以緊緊地抓住石頭的表面。
在生理適應方面,生物演化出應對枯水期惡劣水質的能力。面對枯水期可能出現的高溫和低氧,許多物種表現出更強的廣溫性(eurythermy)和耐低氧能力。例如,生活在下游的鯉魚和鯰魚,相比上游的鮭鱒魚類,能夠在更高溫度和更低溶氧的環境中正常生活。一些極端的例子,如前述的顫蚓和部分搖蚊幼蟲,體內含有高濃度的血紅蛋白,使其能夠在近乎無氧的底泥中生存。還有一些魚類,如鬥魚或部分鯰魚,甚至演化出了輔助呼吸器官(如迷器),可以直接呼吸空氣中的氧氣,從而完全擺脫對水中溶氧的依賴。
然而,最為多樣和靈活的,是生物的行為適應。這是生物應對短期、可預測環境變化的主要手段。面對洪水的來臨,最直接的行為就是尋找避難所(refuge-seeking)。魚類會迅速地從主河道中央,移動到流速較慢的岸邊區域、倒木後方的渦流區,或是湧入洪水淹沒的洪氾平原植被中。許多無脊椎動物則會向河床深處的間隙帶(hyporheic zone)進行垂直遷移,躲入礫石層的深處,以逃避地表最強的水流衝擊。這種利用空間異質性來躲避干擾的行為,是生物得以在洪水中存活的關鍵。
面對枯水期的棲地縮減,生物的行為也極為關鍵。當水位下降,水體變得擁擠時,一些活動能力強的魚類會進行小範圍的遷移,主動尋找並聚集到那些水深、水溫和溶氧條件更好的深潭或有地下水補給的冷水區。這種對優質避難所的利用,是度過枯水期「瓶頸」的核心策略。
更為精妙的是,許多生物的生活史策略(Life History Strategy)與河流的水文情勢實現了完美的同步。它們的繁殖、生長和遷徙等關鍵生命活動,都精確地安排在最有利的時間窗口。例如,許多中下游魚類(如鯉科)的繁殖高峰期,恰好與洪水脈衝的初期相吻合。成魚會利用上漲的洪水,進入被淹沒的洪氾平原產卵。這確保了它們的後代能夠在一個食物最豐富、空間最廣闊、敵害相對較少的「育幼場」中孵化和成長。而當枯水期來臨,這些幼魚已經長得足夠大,能夠游回主河道,應對更嚴酷的競爭環境。
對於許多水生昆-蟲而言,它們的生長和羽化週期也與流量變化緊密相關。一些物種會調整其生長速率,確保在洪水高風險期,自身處於體型較小、更易躲藏的幼蟲階段;而在水流相對穩定的時期,則快速生長。它們的羽化(從水生幼蟲變為陸生成蟲)時間,也常常集中在流量較為穩定的季節,以避免在脆弱的羽化過程中被洪水沖走。
總結來說,河流中的生物並非被動地承受流量的變化,而是主動地、多層次地與之共舞。從身體的形狀,到血液的化學成分,再到遷徙和繁殖的時機,處處都體現了對水文節律的深刻適應。這種長久演化而來的適應性,賦予了河流生態系統強大的韌性。然而,當人類活動(如修建大壩、改變水文情勢)徹底打亂了這種自然的節律,生物原有的適應策略就可能完全失效,從而導致種群的衰退甚至滅絕。保護河流自然的流量變化模式,就是保護其億萬年來演化出的生命智慧。
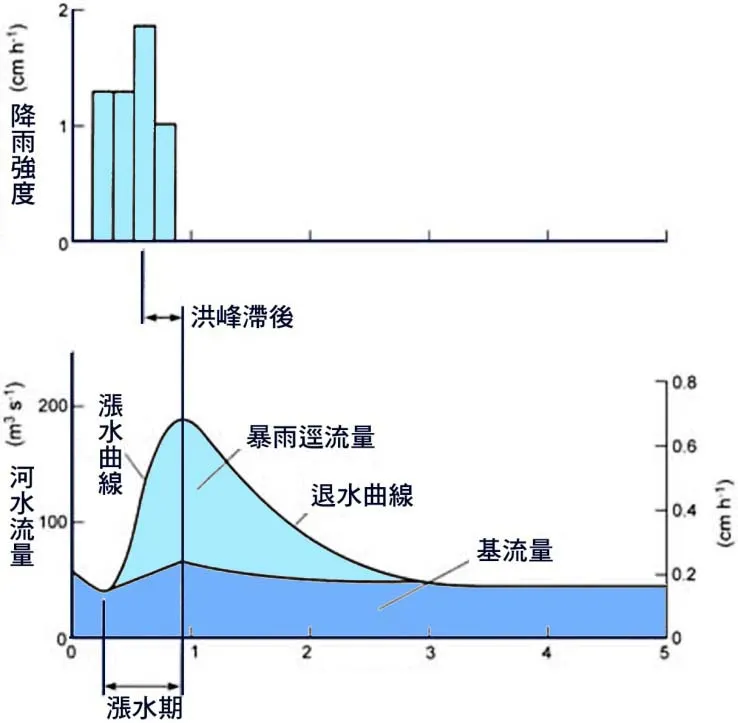
2.4.4 水文週期的生態節律
河流的生命,並非以一種恆定的速率在運轉,而是跟隨著水文週期(Hydrological Cycle)的節奏,呈現出鮮明的生態節律(Ecological Rhythms)。這個節律,以洪水期和枯水期的交替為主要脈絡,貫穿了河流生態系統的方方面面,從初級生產力的波動,到物種的繁殖與遷徙,再到整個群落結構的季節性更替。水文週期就像是河流生態系統的「年曆」和「指揮棒」,規定了各種生態過程上演的時間和順序,使得整個系統呈現出週而復始、充滿動感的生命循環。
初級生產力的節律是整個生態節律的基礎。在溫帶或亞熱帶地區,河流的生產力通常在春末夏初開始飆升。此時,光照增強,水溫回升,而春季的融雪或降雨帶來了較高的流量和來自陸地的營養鹽。附著在石頭上的藻類和水生植物開始快速生長。在中下游的大型河流中,洪水脈衝淹沒了洪氾平原,在廣闊的淺水區觸發了浮游植物的爆發性增長(藻華)。這個生產力的高峰期,為整個食物網提供了豐富的能量基礎。而到了秋冬季節,隨著光照減弱、水溫下降,以及流量的減少,初級生產力也隨之回落到一個較低的水平。
與生產力節律緊密耦合的,是消費者群落的季節性演替。以水生昆-蟲為例,它們的物種組成和豐度在一年中會發生顯著變化。一些物種是「慢季節性」(slow seasonal)的,它們的生命週期長達一年甚至更久,會經歷完整的洪水和枯水週期。另一些則是「快季節性」(fast seasonal)的,它們的生命週期只有幾個月,能夠在水文條件適宜的「窗口期」內,迅速完成從卵到成蟲的發育。因此,在春季,我們可能會看到一群以刮食新生藻類為主的蜉蝣大量出現;到了夏季,隨著水溫升高,另一批更耐高溫的蜻蛉稚蟲可能成為優勢物種;而到了秋季,隨著大量落葉進入溪流,撕食性的石蠅和石蠶蛾的幼蟲則開始活躍起來。這種物種在時間上的交錯出現,實現了對不同季節可利用資源的有效劃分,降低了物種間的競爭。
魚類的生命活動更是嚴格地跟隨著水文週期。如前所述,許多魚類的繁殖活動與洪水期精確同步。除了利用洪氾平原,一些在急流礫石灘產卵的魚類(如鮭鱒魚),它們的繁殖時間也經過精心選擇。它們通常在秋季產卵,此時水溫下降,且即將進入流量較低的冬季,這可以確保魚卵在相對穩定的水流環境下孵化,避免被冬季或早春的洪水沖走。幼魚在春季孵化,恰好能趕上水溫回升、食物(小型無脊椎動物)開始豐富的時期。許多魚類還會進行季節性的棲地轉換。在洪水期,它們可能會分散到廣闊的洪氾平原或岸邊新淹沒的區域覓食;而在枯水期,它們則會集中到深潭等避難所中。這種跟隨水位的「漲出落歸」,是它們利用整個河流-洪氾平原生態系統的關鍵策略。
整個生態系統的物質循環也呈現出明顯的節律性。洪水期是物質橫向交換的高峰期,營養鹽和有機物在主河道和洪氾平原之間進行大規模的雙向流動。同時,洪水也是縱向輸送的高峰期,大量的沉積物和有機質被從上游輸送到下游。而枯水期,則是物質內部再循環和處理的關鍵時期。在緩慢的水流中,有機物有更長的時間沉降和被分解,營養鹽在局部被吸收和再利用。河流的自淨作用,在這兩個時期表現出不同的特點:洪水期以物理的「稀釋和沖刷」為主,而枯水期則更依賴生物和化學的「分解和吸收」。
總結來說,水文週期為河流生態系統注入了一種深刻的時間秩序。洪水與枯水,如同樂章中的高潮與間奏,共同譜寫了河流生命的交響曲。每一個物種,每一個生態過程,都在這個宏大的節律中找到了自己的位置。這種生態節律的存在,賦予了河流生態系統以可預測性和穩定性。然而,這種穩定性是建立在水文週期的可預測性之上的。當全球氣候變化導致降雨模式改變(如更頻繁的極端乾旱和極端洪水),或當人類工程(如大壩)徹底改變了河流的脈動時,這個千萬年來演化形成的生態節律就會被打破。生物的生活史策略與錯位的環境不再匹配,生態過程發生紊亂,整個河流生態系統的健康和穩定性將面臨嚴峻的挑戰。保護河流,不僅要保護其空間的完整,更要保護其時間上的自然節律。